水稻半卷葉控制基因SRL9及其葉形改良的用途的制作方法
本發明屬于植物基因工程領域。具體地說,本發明涉及一種利用半卷葉突變體圖位克隆獲得水稻半卷葉控制srl9基因,以及利用轉基因互補實驗確認該基因的功能;同時還涉及利用該基因開展分子設計育種,塑造水稻理想株型,提高農作物的產量。
背景技術:
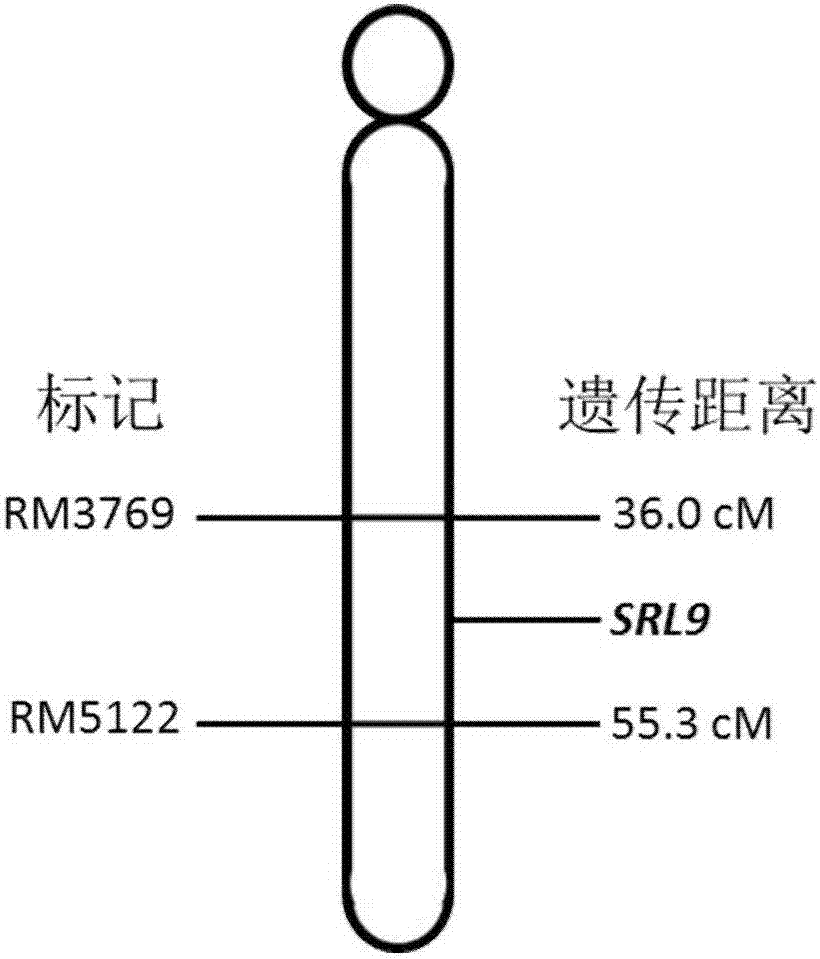
:水稻是單子葉模式植物,其葉片的外部形態結構由葉片、葉鞘、葉枕以及葉舌和葉耳組成。其中葉片包括表皮、葉肉和葉脈三個部分。葉片極性發育決定葉長、葉寬、葉片卷曲度等葉片形態建成的主要指標。水稻葉片為等面葉,葉片角質層、各級維管束數目和結構、葉肉細胞、近軸面泡狀細胞及遠軸面厚壁組織等都可能導致正常葉片的極性改變。近年來,科學家們在分子水平對葉片形態建成調控機制開展研究,分離與克隆了多個與葉片形態發育調控相關的基因。自2009年zhang等借助水稻葉形突變體正向克隆第一個卷葉基因sll1以來,科學家們已先后完成了20余個與葉片形態發育相關基因的克隆及調控機制的初步研究。植物葉片的適度卷曲,尤其是最上面三片功能葉的適度卷曲,會使葉片直立上沖,增大整個群體的受光面積,不遮陰,使下部葉片也能夠有光能吸收,從而提高光能產物的吸收,提高產量潛力。大部分卷葉基因與近軸面泡狀細胞的發育調控相關,其數目和大小的改變導致葉片出現正卷或反卷;acl1基因突變,其突變體的葉片近軸面泡狀細胞的數目和大小增加,使表皮細胞擴張,導致植株的葉片向外卷曲;roc5,編碼一個hd-zipiv基因家族成員(第四類亮氨酸拉鏈同源框家族),該基因的過表達植株的葉片上表皮泡狀細胞的數目和大小都有所減少,造成內卷的表型;而共抑制植株的葉片泡狀細胞數目和大小卻都有所增加,導致葉片反卷,也說明roc5基因具有保守性。rl14,編碼一個2og-fe(ii)氧化酶家族的蛋白,其主要在葉片維管束周邊的葉肉細胞中積累表達,葉片近軸面的泡狀細胞失水皺縮,且葉片中纖維素和木質素的含量也發生了顯著的變化,表明可能是rl14通過影響葉片次生細胞壁的組分含量而影響葉片的水分運輸,使葉片泡狀細胞的形態發生異常,最終導致植株的葉片向近軸面卷曲。srl1編碼一個糖基磷脂酰肌醇錨定蛋白,通過負向調節編碼液泡h+-atpase亞基和h+-焦磷酸酶基因的表達,進而抑制葉片中泡狀細胞形成,調控葉片卷曲。oszhd1,編碼一個鋅指同源結構域類同源盒轉錄因子,過表達植株的葉片近軸面泡狀細胞數目增加,并且排列異常,導致葉片向遠軸面卷曲,造成葉片反卷的表型出現。myb103l,編碼一個r2r3型myb轉錄因子,過表達植株的葉片泡狀細胞變小,導致葉片正卷,且體內的纖維素合成酶相關基因的表達量受到影響,且纖維素含量增加,而干涉植株的葉片纖維素含量和機械強度都有所降低。sll2定位在第7染色體上,該基因的突變導致從6葉期開始出現內卷,伴隨著光合作用的增強及株高和分蘗數目的減少,其組織細胞學分析表明,泡狀細胞的缺收縮是導致葉片內卷的主要原因。rel1基因編碼的蛋白,能夠通過協調油菜素內酯信號調控葉片的形態,rel1突變體中泡狀細胞數目和大小都增加,導致葉片反卷。rel2基因編碼一個包含duf630和duf632結構域的功能未知蛋白,rel2近軸面表皮的泡狀細胞數目增加,中部泡狀細胞減小,導致葉片正卷。還有一部分基因,通過調控近-遠軸面不同組織的發育,影響葉片卷曲度,包括sll1、adl1、osago7、srl2等。其中,sll1編碼一個屬于kanadi家族shaqkyf類myb轉錄因子,該基因的突變導致葉片遠軸面的厚壁組織細胞程序性死亡,不能形成厚壁細胞,導致葉片極端的向近軸面卷曲。adl1編碼一個類鈣蛋白酶半胱氨酸蛋白酶,該基因突變導致遠軸面的葉肉細胞和上皮細胞近軸化,分化成類泡狀細胞,使葉片向遠軸面卷曲,是維持葉片近遠軸發育的重要基因。osago7編碼一個屬于argonaute(ago)蛋白,控制葉片近-遠軸面的發育,使葉片向上卷曲,增強了葉片的直立性。srl2基因主要在葉片的維管束、葉鞘、根中表達,特別是在它們的厚壁細胞中表達,通過調控遠軸面厚壁組織的分化,使遠軸面近軸化,導致葉片正卷。此外,仍有一小部分基因,如cfl1(通過調控細胞質的發育而影響葉片卷曲。技術實現要素:本發明要解決的技術問題是提供一種ems誘變獲得水稻半卷葉突變體中克隆的新半卷葉基因srl9,屬于kanadi家族myb轉錄因子,其通過調控葉片近軸面泡狀細胞和維管束發育,調節葉片卷曲程度。為了解決上述技術問題,本發明提供一種水稻半卷葉基因srl9編碼的蛋白質,該蛋白質具有seqidno:2所示的氨基酸序列。作為本發明的水稻葉形控制基因srl9編碼的蛋白質的改進:氨基酸序列還包括在seqidno:2所示的氨基酸序列中添加、取代、插入或缺失一個或多個氨基酸或其他物種中的同源序列而生成的氨基酸序列或衍生物。本發明還提供了編碼上述蛋白質的基因,該基因具有seqidno:1所示的核苷酸序列。作為本發明的基因的改進:核苷酸序列還包括在seqidno:1所示的核苷酸序列中添加、取代、插入或缺失一個或多個核苷酸而生成的突變體、等位基因或衍生物。本發明還提供了含有上述基因的質粒。本發明還提供了含有上述基因的植物表達載體。本發明還提供了一種宿主細胞,該宿主細胞含有基因序列,該細胞為大腸桿菌細胞、農桿菌細胞或植物細胞。本發明還提供了上述基因的用途---用于改良水稻葉形,用雜交技術(常規雜交技術)構建基因替換的水稻,所述基因替換水稻的葉形得以改良,水稻葉片發生卷曲。所述基因替換水稻的結實率和產量得以提高。具體的說:本發明所提供的從水稻突變體中克隆的半卷葉新基因srl9,具有seqidno:1所示的核苷酸序列。也包括在取代一個核苷酸而產生的突變體等位基因,還含具有相同功能并能達到本發明目的的基因序列。本發明中seqidno:2所示的蛋白質,還包括其中進行一個或幾個氨基酸的替換、插入或缺失氨基酸所獲得的功能類似物。本發明所提供了含有seqidno:1所示序列的基因或含部分該基因片段的載體,如圖4所示,該載體可以表達由上述核苷酸序列編碼的多肽或同源類似物。本發明利用ems誘變獲得的半卷葉突變體,通過圖位克隆得到具突變位點的半卷葉控制基因srl9,最終驗證基因功能。此發明有助于闡明水稻葉形建成的分子機理,并通過分子設計育種改良葉形,為理想株型高產育種奠定理論和材料基礎。實現本發明的具體技術步驟如下:一、水稻卷葉突變體srl9的分離和遺傳分析本發明所采用的水稻半卷葉突變體(semi-rolledleaf9,srl9)是將秈稻品種“蜀恢527”經1%(w/v)濃度的化學誘變劑(ethylmethanesulphonate,ems)處理后,再通過大量篩選而得到的表型穩定的半卷葉突變體。該突變體srl9葉片具半卷特性,其如圖1所示。雜交之后產生f1代自交產生的f2代群體,f2代群體共4319株,其中正常葉為3274株,卷葉為1045株,經卡平方測驗,正常葉與半卷葉個體的分離比符合3:1,表明我們所得到的是一個符合單基因控制遺傳規律的半卷葉隱性突變體。二、圖位克隆控制水稻葉形的srl9基因1、srl9基因的初步定位為了分離srl9基因,本發明首先建立了一個大的多態性高的定位群體,由srl9與粳稻品種日本晴(nipponbare)雜交后自交而形成的f2群體,再通過圖位克隆的方法,并利用sts、ssr等分子標記對srl9位點進行初步定位,將其初步定位在第9染色體的長臂上,并介于rm3769和rm5122兩標記之間。2、srl9基因的精細定位通過對bac克隆ap005904的序列分析,發展了3個新的ssr標記和15個sts標記,如表1所示,將srl9精確定位于sts(sequencetaggedsite)標記sr9-5904-18(f:gtcatcagtcaccgtgttca;r:catgcggaatctgtaacttg)和ssr分子標記sr9-5904-s3(f:cgtcagcctacacatcatctc;r:cttgagattggtttgcatcat)之間,13.834kb的范圍之內。通過測序分析此區段的開放閱讀框(orf)推測候選基因,將srl9基因精確定位于bac克隆ap005904的103061-108515的位置(圖3)。3、srl9基因的鑒定和功能分析利用pcambia1300質粒構建srl9互補載體。以蜀恢527的基因組dna為模版,用序列為5'-cgagatcttagattatttttattcgtatataag-3'和5'-ggcctgcagggattaaagaaagaagc-3'的引物進行pcr擴增,獲得的7989bp片段pcr擴增產物用bglii和sdai進行完全雙酶切,得到全長為7980bp的基因組dna片段,包含srl9基因的啟動區的1810bp,5455bp的編碼區,715bp的srl9基因的下游序列,將該片段后連到pcambia1300載體多克隆位點bamhi和psti之間獲得pcambia1300-srl9互補載體(bglii與bamhi,sdai與psti分別為同尾酶)(圖4)。通過轉基因技術,進行功能互補的轉基因研究,結果表明本發明獲得了使突變體恢復正常葉片形態的轉基因水稻(圖5),證明了本發明正確克隆了srl9基因,明確srl9基因的dna序列(seqidno:1)和cdna序列(seqidno:3),氨基酸序列分析表明srl9基因編碼kanadi基因家族myb轉錄因子(seqidno:2)。綜上所述,本發明從水稻半卷葉突變體(semi-rolledleaf9,srl9)中克隆并鑒定了控制水稻葉片卷曲程度的基因,將其命名為srl9,并通過互補實驗進行基因功能驗證。在水稻中,平展葉片的srl9基因單堿基替換后編碼的氨基酸產生變化,基因功能導致葉片近軸面泡狀細胞發育異常,造成半卷曲的表型。圖位克隆的結果表明,該基因編碼的myb轉錄因子,通過調控近軸面泡狀細胞和遠軸面葉肉細胞發育,控制水稻葉片卷曲度。srl9基因單堿基突變后,葉片中泡狀細胞數目增多,導致葉片呈半正卷狀態,改變了平展野生型平展葉片的受光姿態,葉片在相同光照情況下,可以增加受光面積,提高光合作用效率,增加糧食產量。葉形作為作物重要的農藝性狀,是理想株型的重要組成部分,在品種改良和分子設計輔助育種中占有重要的地位。而適度卷曲的葉片是水稻理想株型的重要內容之一,改良植物葉片的卷曲程度,從而提高光合作用效率,加速干物質的積累,提高作物產量,具有廣闊的應用前景。該發明可以為水稻株型改良和分子設計育種提供新的基因資源和理論指導。附圖說明下面結合附圖對本發明的具體實施方式作進一步詳細說明。圖1是水稻苗期半卷葉突變體srl9和野生型的表型圖;圖2是半卷葉基因srl9在水稻第9染色體長臂上的初定位圖;圖3是半卷葉基因srl9在水稻第9染色體上的精細定位圖;圖4是互補載體pcambia1300-srl9質粒圖譜;圖5是功能互補實驗t2代轉基因互補植株表型圖;圖6是利用半卷葉基因srl9改良日本晴葉形技術路線圖。具體實施方式下面結合具體實施例對本發明進行進一步描述,但本發明的保護范圍并不僅限于此:實施例1:水稻半卷葉調控基因srl9的克隆1、水稻材料水稻(oryzasatival.)突變體srl9(semi-rolledleaf9,srl9),其原始野生型材料為秈稻品種“蜀恢527”。水稻(oryzasatival.)突變體srl9是將秈稻品種“蜀恢527”經1%(v/v)濃度的化學誘變劑(ethylmethanesulphonate,ems)處理后,再通過大量篩選而得到的表型穩定的突變體srl9。該突變體除了葉片呈半卷曲外,其它表型均與野生型相似,如圖1所示。從圖1中,我們可以看出,在水稻的苗期,突變體的葉片呈現半卷曲的狀態。2、分析和定位群體純合的半卷葉突變體srl9與粳稻常規品種日本晴(nipponbare)進行雜交,f1代自交,得到f2群體,f2代群體共4319株,其中正常葉為3274株,卷葉為1045株。選擇該1045株半卷葉突變體srl9表型植株作為定位群體。在分蘗期每株取1克左右的嫩葉,用來提取總dna。3、ssr和sts標記定位srl9基因采用改良的ctab法提取水稻葉片用于基因定位的基因組dna。取3-5cm水稻葉片,剪碎放入2.0mleppendorf離心管中,加入1粒鋼珠,700ul提取液,置于tissuelyseriisystem組織研磨器,研磨2-3min,65℃恒溫水浴40-60min,再用氯仿萃取,吸取上清液,加入等量冰凍的異丙醇,離心,用70%酒精洗滌,晾干即可加入150μl超純水作為dna樣品。每一個pcr反應用2-3μldna樣品。srl9基因的初步定位:在srl9與日本晴(nipponbare)組合的f2群體中隨機選取1045卷葉株中的186株,組成的小群體進行ssr分析,根據公布的粳稻和秈稻創建的分子遺傳圖譜,選取近似均勻分布于各條染色體上的ssr引物,利用mjresearch的ptc-200型梯度熱循環儀進行pcr擴增,pcr擴增反應總體積為20μl,其中包括50mmol/lkcl,10mmol/ltris-cl(ph9.0),1.5mmol/lmgcl2,200μmol/ldntp,50-100ng的基因組dna,1個單位的taq聚合酶和0.1μmol/l引物,其余為蒸餾水。擴增程序為94℃預變性4min,進入循環擴增:94℃變性30s,各引物及其退火溫度見表1,退火時間30s,72℃延伸40s,循環40次;72℃延伸10min。經5%瓊脂糖凝膠電泳分離和溴化乙啶(eb)染色,檢測pcr產物的多態性。最終將srl9初步定位在第9號染色體長臂rm3769和rm5122兩分子標記之間。srl9基因的精細定位:在srl9與日本晴(niponbare)組合的f2群體中共選取1045株(包括初定位的186株)隱性個體,在初定位的基礎上繼續設計sts標記,通過對秈稻品種9311和粳稻品種日本晴進行序列比對分析,發展了18個新的具有多態分子標記(表1),其中ssr(simplesequencerepeats)分子標記3對,具體為sr9-5904-s1、sr9-5904-s3和sr9-5904-s6;sts(sequencetaggedsite)分子標記15對,具體為sr9-5321、sr9-5556、sr9-6562、sr9-6453、sr9-5904-2、sr9-5904-5、sr9-5904-8、sr9-5904-10、sr9-5904-11、sr9-5904-12、sr9-5904-13、sr9-5904-18、sr9-5904-24、sr9-5577和sr9-5421。利用mjresearch的ptc-200型梯度熱循環儀進行pcr擴增,pcr擴增反應總體積為20μl,其中包括50mmol/lkcl,10mmol/ltris-cl(ph9.0),1.5mmol/lmgcl2,200μmol/ldntp,50-100ng的基因組dna,1個單位的taq聚合酶和0.1μmol/l引物,其余為蒸餾水。擴增程序為94℃預變性5min,進入循環擴增:94℃變性45s,各引物及其退火溫度見表1,退火時間40s,72℃延伸50s,循環35-39次;72℃延伸10min,冷卻降溫至16℃后備用。經4%-5%瓊脂糖凝膠電泳分離和gelred染色,檢測pcr產物的多態性。最終將srl9精確定位于ap005904的bac上,位于sts(sequencetaggedsite)標記sr9-5904-18和ssr分子標記sr9-5904-s3之間共13.834kb的范圍之內。基因定位的分子標記序列如表1所示。表1、用于基因定位的分子標記序列4、srl9基因預測、cdna全長基因的獲得與功能的預測:根據精細定位的結果,在13.834kb范圍內根據riceautomatedannotationsystem(http://ricegaas.dna.affrc.go.jp)的預測,發現在此區間內只含有1個候選基因,我們設計了這個候選基因的測序引物(見表2),采用pcr方法分別從srtl9和野生型秈稻品種蜀恢527基因組中擴增出候選基因片段,委托上海英駿生物技術有限公司進行測序(pcr擴增反應體系和擴增程序參照上文)。分析發現蜀恢527的srl9基因的dna片段中,從起始密碼子atg開始的第8個堿基產生了有c到t的替換,導致編碼的氨基酸密碼子由ccg(pro,脯氨酸)到ctg(leu,亮氨酸)的改變。根據bac克隆ap005904序列的基因注釋信息(ncbi),預測該基因編碼kanadi基因家族myb轉錄因子,通過調控葉片近軸面泡狀細胞和維管束發育,調節葉片的卷曲程度。經過pcr擴增得到了srl9基因的cdna全長序列(seqidno:3)。srl9基因的核苷酸序列如seqidno:1所示。該水稻半卷葉基因srl9編碼的蛋白質具有seqidno:2所示的氨基酸序列。seqidno:2中的“*”表示終止密碼子(tag)。表2、用于候選基因測序的引物及其pcr退火溫度實施例2:植物轉化利用mjresearch的ptc-200型梯度熱循環儀進行pcr擴增。以蜀恢527基因組dna為模版,用序列為5'-cgagatcttagattatttttattcgtatataag-3'和5'-ggcctgcagggattaaagaaagaagc-3'的引物pcr進行擴增。pcr擴增反應體系:總體積為40μl,其中包括50mmol/lkcl,10mmol/ltris-cl(ph9.0),1.5mmol/lmgcl2,200μmol/ldntp,50-100ng的基因組dna,5個單位的taq聚合酶和0.1μmol/l引物,其余為蒸餾水。擴增程序:94℃預變性4min,進入循環擴增;94℃變性50s,退火溫度60℃,退火時間40s,72℃延伸4min,循環35次;72℃延伸15min,降溫冷卻至16℃后備用。pcr擴增獲得的7989bp長度的產物,用bglii和sdai進行完全雙酶切,得到全長為7980bp的基因組dna片段,包含srl9基因的啟動區的1810bp,5455bp的編碼區,715bp的srl9基因的下游序列,將該片段后連到pcambia1300載體多克隆位點bamhi和psti之間獲得pcambia1300-srl9互補載體(bglii與bamhi,sdai與psti分別為同尾酶)。這個質粒通過電擊的方法轉入農桿菌(agrobacteriumtumefaciens)株系eha105中轉化水稻。我們利用突變體srl9成熟胚誘導的愈傷組織,經過誘導培養基培養2周后,挑選生長旺盛的愈傷用作轉化的受體。用含有雙元質粒載體(pcambia1300-srl9)的eha105菌株侵染水稻愈傷,在黑暗、25℃條件下共培養3天后,在含有50mg/lhygromycin的篩選培養基上光照培養14天左右(光照強度為13200lx,溫度為32℃)。將預分化的愈傷轉至分化培養基上在光照條件下(光照強度為13200lx,溫度為32℃)培養一個月左右得到抗性轉基因植株。對植株進行鑒定和連續的觀察,發現植株葉片恢復了正常的形態,與同一生長階段的突變體比較,葉片不再卷曲。如圖5所示。實施例3:改良水稻葉形的方法利用半卷葉突變基因srl9改良日本晴平展葉片卷曲度,改良方法如圖6所示。具體步驟為:以半卷葉突變體srl9為半卷葉調控基因的供體親本,以平展葉的粳稻日本晴為半卷葉基因srl9的受體親本,雜交后,用日本晴作為輪回親本連續回交4代再自交2代的基礎上,用sr9-5904-18和sr9-5904-s3兩個分子標記(兩標記之間只含有一個srl9基因),篩選含有供體突變體srl9中的突變基因srl1片段株系;結合株型,篩選接近日本晴的半卷葉植株,繼續與日本晴回交1代并自交,在其后代中用分子標記sr9-5904-18和sr9-5904-s3篩選含有半卷葉突變基因srl9片段且表現為純合的日本晴近等基因系,用于農藝性狀和產量相關性狀調查。通過對篩選到的5個葉形改良后的半卷葉日本晴近等基因系(neariso-geniclines,nil)植株與日本晴株高和生育期的考察發現,兩者之間沒有顯著差異(表3);從產量性狀比對結果看,雖然半卷葉日本晴近等基因系的平均千粒重相對日本晴有一定程度增加,但兩者之間差異不顯著;但從結實率和每畝產量考察結果來看,除了srl9-npb-nil1和srl9-npb-nil5兩個株系以外,其余所有葉形改良后的半卷葉日本晴近等基因系株系(srl9-npb-nil2~4)的平均結實率要顯著高于原受體親本株系日本晴;而所有半卷葉日本晴的近等基因系(srl9-npb-nil1~5)的每畝產量相對日本晴都顯著增加(表4)。說明導入半卷葉調控基因srl9的葉形改良日本晴能顯著增加水稻產量,srl9基因在水稻葉形改良和產量育種的實踐中具有重要的實際應用意義。表3受體親本日本晴及葉形改良的日本晴半卷葉近等基因系(nil)株高和生育期評價株系名稱全生育期(天)株高(cm)日本晴(npb)104.6±0.5889.4±4.61srl9-npb-nil1105.3±0.5888.6±3.65srl9-npb-nil2103.6±1.5389.8±4.49srl9-npb-nil3104.6±0.8788.7±3.36srl9-npb-nil4105.3±0.4887.9±5.08srl9-npb-nil5104.6±1.5688.4±3.65表4受體親本日本晴及葉形改良的日本晴半卷葉近等基因系(nil)產量性狀調查最后,還需要注意的是,以上列舉的僅是本發明的若干個具體實施例。顯然,本發明不限于以上實施例,還可以有許多變形。本領域的普通技術人員能從本發明公開的內容直接導出或聯想到的所有變形,均應認為是本發明的保護范圍。<110>中國水稻研究所<120>水稻半卷葉控制基因srl9及其葉形改良的用途<160>3<210>1<211>5455<212>gdna<213>水稻(oryzasativa)<400>1atggcgctgatgatgctgcagccgctcgacgccggcggcggcgcgtcggcgccgccgccg60ccgatccgcgggatacctatctacaacggccccggcgggttcccgttcctgcagccgtcg120cccaccgccggcgacgtcggccaccaccaccaccaccaccccaagatgggattctacagc180tcgtaccaccacccatccacgtggccctccacgtcgccgtccccgctcgcggcgccgccg240ggcgccgcgtcgtcgccgctcgaccccacggcggcgttcctctcctccccccaccaccgg300atgctgtccgccgcctcggggaggctcaacggcatgctctccgtctccgacaccctccgc360agctacggcgtccccggcgccgccgcccccggcgtcatcggcggcgcgcaccaccaccac420caccacctccacggcggccagccgttcgtcggcgccctcgcgtcccgcttcatgcccaag480ctccccgccaagcgcagcatgcgcgcgccgcgcatgcgctggacgagcaccctccacgcc540cgcttcgtccacgccgtcgagctcctcggcggccacgagagtacgcccccgccatcatcg600tcgtcgtcctcctcctccttctcaccaccatcatcaacgagctcgatcaatctgatgaga660atccaatcttgttcttggcaggggcgacgcccaagtcggtgctggagctcatggacgtca720aggatctgacgctagcgcatgtcaagagccacctccaggtatcccgccattgccgaccaa780ttctcacagctcgatcgatcgatcaaacaccactcaccactgcaccatctctctctttct840ctctctcgatttcgattccattcagcttctgttcttgatcatcttgtttgggtgagacaa900agatgccaatgcagtaactcctgagttagtgaagaaactgccattcttgggagcagagag960agagagtgtgtgtgtgacatgtttggggatgtgtgtgcaagagagaagagagagagagag1020gggaaaaaagttgccatcattacacaataatgcattgcatctgcatctgtgctactagct1080ccttccttagctctagtcatactagcgtatatgtgcacggcccaaagcactctcacacac1140aaacacaagagagagagagagagagagaagaagaagatgagagatgagagagagtaggac1200atattttgtgtgtgtttctttgctgcttgtcttttgcaatggcttcaacctcctggtttc1260attttgatctcacaaaaaatgtgtgtgtgtgtgtaaatatatatattgatcttctttttg1320ttggacttttgtgttttattactgcagatgtatcgcaccgtgaagagcactgacaagcct1380gcagcctcttcaggtgattttttttaccatggcagctactagtagtagtagtagtagcta1440atttagctaagctccattttgcttttctttttttttggcgttgtgcattgcgttttacat1500tacattgttgctatggtttttttgacgtgtagggccggcggacggcggctccggcgacga1560ggagttcgccggcggcgggcaggcggcgtcgggcggcggcgacagcatgtgcctgagggg1620tggcggcggcggcggggtggccgcggcggcgttcgcggagcacggccggtcggcgtcgga1680gggcgccgccagctcggtcggcggcggcggcggcggcgacatggaccagtcgtcggccgg1740caacaccagcaccaccaggtggagcaactcctcaaggtacacacatggcagagttgcatt1800tcaagtctttcagattaattatatatctacattgttatgccatctttaattaattaaaaa1860tgaaattaatcaatcagaatgtcatgagatgatgatgatgatgataataatactaggata1920ggaagaggatgatatgatggatgggtgtgctatgtgtgtttttaagtgtgttttagctta1980gaaccaaaggcaacaccaacaaactttggcatgcattgatgatgtatctaaatatgagtt2040agtgacatgcaaaattaacctgccaagtacaacctctatttattaattcccaaaaaaatc2100tcatggaggagaagggagagagagagatagtagtaatgtgtggtgatgcatggctaaaaa2160agcttagcaaaagaatgttttgggggcacacacacacacacctggagagggagttgccta2220tgtttaggtggccaatgttgttatgtcacacaaaagaccaaaaagtcacacaccccatgg2280cttggcctcctcctccccccttgacctgtgcagttagctaggcttcttctcttctctccc2340tctgtgtttggtttttgtggagctaaaaacattttaaaaatcttgcaaacattgtgtgta2400aaaataggttcttgggctctgcccattgtcctctagcttctctcaaatctctctctctct2460ctctctctctctctctctctttctcaaacctagggaggggagggttactcaggcagggct2520ccattatcatttcttatcaatgcatggctctgattccagggggatggggctttgacctga2580catgccctattgcctaggaatatgtaaccaaggacttgctgcattcttggttctttctcc2640ctctctcaccagtctcttcaatgcccttttcttccacctcttgggcaattattagcatgg2700atggagctcaccttttcactctaccctctagctgtgattaaatcattgactaatcctaat2760tcataaatataagggattcaactatgtaactagatgtactagatcctagtattactagtg2820tgttttgattctgttaggaggagtgattagcgtttgtcgtaagtactagtattaaactat2880aggggattagtgtagatcatttttttaaaaattagtgagtgtgcttgggcagattttact2940ccagccattggcgtgaaaactttgatctttgggcagcttggaagcaagtacagttcaacc3000tgtccttggctgcattcgctgctttttctttctcaggactcactgcaactttcttgccat3060ccccaacacctccctttcggtcgtgtttgatcagttgcattgctggtttcttcgccgctt3120taatttctccatctttcttctgttccttattggctaagtaaaaaatcattctcagcaatc3180aagttttgaaattcaaattagcttcaggttttatctatctaataattctctgactagttc3240tctttctttcaagacctgttcttgttcagaactcttatgcagttgagagttcagttgttc3300atggaagtaatggcaatgcctggtctagtctttgagcagtatcctccttatgctcttgat3360tttccttcactttagccgtttcagtcaaactttggggacaaattaaagtcagctggtcaa3420gtgcatacataccacgtcatattttttagatgaatccattgtagtattaaatactagtga3480caaaaaaatataactaaatggcattaaattgcaaccttcttccccaaaataattgggcac3540aactaccttgcttgcatatgctttatagtactcctacttattacttccttgaatccatga3600ggattccaggaatttggtcaggtcaaatgtccaggaggaattaggctatctttttttatc3660ttcttctttttcttctaggtagtaggatcgttgtgcacatataaacaaaacaaaatagaa3720atttaagaggaaaccctcccaaaaaaaatacgactatcaattatgagtgattttacagtt3780cttgaggaggtacaatgatacattttctattttaaaattcagtaactttgatactatgag3840gtacagaaatttgcacaaaaaaaatttggtacctctcaagtacttaaaatatcttatcaa3900ttatatttcagcaatgttgtcgttgctcgagttgtgcaaaaacatttctgatgaacaagt3960ggagatgtatgcatatttagagagggaaacacatgcattttttaatatacattctcatga4020caaattttggtgttctcgtctcattttagtctagcttagattaatggcatccatatattg4080gtgaactgaaacagattgtatatgtccagtcttgaacaaacttcattatacaatgcataa4140tcagaaaagatacagacatatattggcagatcacataaagtggtgtatctatgattgggt4200ggtattttggacatacaaagtctactggtccctcatttgttaacagcatacatttgttcc4260tatcaagacaaaagcactttacaacagtttggtagtaagttggtgttatgtgcgtgtcta4320tttagatagagtggtagtaaaggaaggacactttttcttgatataccaaaatcacacata4380aaaaagaagtgtcattttcatgatcaacatgtggggtatgaacaaaaatgttctcccccc4440tccaaccaacattgaagtcatcacacataaaaagcaaaaacatttaacaataatacatgc4500agacacatgtgcatgcatgtgtggatcacgcaaggacatactaattcaattaggtattca4560caattaattagtgcaggaaatgacatgtaattacaattatatatgcacaacgatccttgt4620gtatttgtgtgacccattgtagtagcatttgattcctttcttgtgcctttctctacattg4680aattcgatctttgccaacagcggcttgctgatgagctcctgtgccattgcactgctgagc4740tgcaccaaacagggacatcttttaagagcttgttgcctggaaatgatccatgtaaccaac4800tgttcaggaagagcacactgtttgctgctcactttagctaaagcaaaactgtgtgcatta4860tgcaatgcatatgcatccttccatctacccctgtgtcctttgccccccaaactgtgtaac4920tacagtttaccatgcacatacatacaatgcacatctacccaattgtagcattgttgcaag4980gcctacttcagaattcagttagtccccaattgatccaactgcacacatatacatttacat5040atacatatacacaaacataccagcttggagaaaaggttgtaacaactttgctaaatgtga5100gatgtgtaatgtgtgttcttcctcttttcagggacccatggctgtcgtccaattcttgca5160acatggacgcccatcgctccgtaggattgtcttctcctattgaggtatatatccttacat5220tgttcaaagcttctttcgattcctagctaactgcatatccatgcagaaattcattagctc5280gcattcattctgaacatgatgtctgaaactctgaattttttttcgtttgtgcgtgtttca5340gaacttggaaccgtgcagatcgagcagctcgcaggtgtccaaccatgagctgagtagccc5400tagtctcgagttcactctagggaggcctgactggcacggtgcagatcatgattag5455<210>2<211>377<212>prt<213>水稻(oryzasativa)<400>2metalaleumetmetleuglnproleuaspalaglyglyglyalaseralapropropro15101520proileargglyileproiletyrasnglyproglyglyphepropheleuglnproser25303540prothralaglyaspvalglyhishishishishishisprolysmetglyphetyrser45505560sertyrhishisproserthrtrpproserthrserproserproleualaalapropro65707580glyalaalaserserproleuaspprothralaalapheleuserserprohishisarg859095100metleuseralaalaserglyargleuasnglymetleuservalseraspthrleuarg105110115120sertyrglyvalproglyalaalaalaproglyvalileglyglyalahishishishis125130135140hishisleuhisglyglyglnprophevalglyalaleualaserargphemetprolys145150155160leuproalalysargsermetargalaproargmetargtrpthrserthrleuhisala165170175180argphevalhisalavalgluleuleuglyglyhisgluargalathrprolysserval185190195200leugluleumetaspvallysaspleuthrleualahisvallysserhisleuglnmet205210215220tyrargthrvallysserthrasplysproalaalaserserglyproalaaspglygly225230235240serglyaspglugluphealaglyglyglyglnalaalaserglyglyglyaspsermet245250255260cysleuargglyglyglyglyglyglyvalalaalaalaalaphealagluhisglyarg265270275280seralasergluglyalaalaserservalglyglyglyglyglyglyaspmetaspgln285290295300serseralaglyasnthrserthrthrargtrpserasnserserargaspprotrpleu305310315320serserasnsercysasnmetaspalahisargservalglyleuserserproileglu325330335340asnleugluprocysargserserserserglnvalserasnhisgluleuserserpro345350355360serleugluphethrleuglyargproasptrphisglyalaasphisasp*365370375<210>3<211>1134<212>cdna<213>水稻(oryzasativa)<400>3atggcgctgatgatgctgcagccgctcgacgccggcggcggcgcgtcggcgccgccgccg60ccgatccgcgggatacctatctacaacggccccggcgggttcccgttcctgcagccgtcg120cccaccgccggcgacgtcggccaccaccaccaccaccaccccaagatgggattctacagc180tcgtaccaccacccatccacgtggccctccacgtcgccgtccccgctcgcggcgccgccg240ggcgccgcgtcgtcgccgctcgaccccacggcggcgttcctctcctccccccaccaccgg300atgctgtccgccgcctcggggaggctcaacggcatgctctccgtctccgacaccctccgc360agctacggcgtccccggcgccgccgcccccggcgtcatcggcggcgcgcaccaccaccac420caccacctccacggcggccagccgttcgtcggcgccctcgcgtcccgcttcatgcccaag480ctccccgccaagcgcagcatgcgcgcgccgcgcatgcgctggacgagcaccctccacgcc540cgcttcgtccacgccgtcgagctcctcggcggccacgagagggcgacgcccaagtcggtg600ctggagctcatggacgtcaaggatctgacgctagcgcatgtcaagagccacctccagatg660tatcgcaccgtgaagagcactgacaagcctgcagcctcttcagggccggcggacggcggc720tccggcgacgaggagttcgccggcggcgggcaggcggcgtcgggcggcggcgacagcatg780tgcctgaggggtggcggcggcggcggggtggccgcggcggcgttcgcggagcacggccgg840tcggcgtcggagggcgccgccagctcggtcggcggcggcggcggcggcgacatggaccag900tcgtcggccggcaacaccagcaccaccaggtggagcaactcctcaagggacccatggctg960tcgtccaattcttgcaacatggacgcccatcgctccgtaggattgtcttctcctattgag1020aacttggaaccgtgcagatcgagcagctcgcaggtgtccaaccatgagctgagtagccct1080agtctcgagttcactctagggaggcctgactggcacggtgcagatcatgattag1134當前第1頁12
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1