桑樹乙烯響應因子及其應用的制作方法
本發明涉及植物基因工程領域,尤其涉及一種桑樹乙烯響應因子(mmerf)基因的克隆和其在脅迫條件下的表達。
背景技術:
植物在生長發育過程中會遇到低溫、干旱和鹽堿等一些非生物性脅迫,它們是目前影響植物生長及作物產量的主要限制因子,植物為了適應周圍的環境,當受到低溫、干旱、高鹽等脅迫時,細胞迅速感知外界脅迫信號并通過一系列信號傳遞過程,產生應激信號并匯集到細胞核內,最終激活特定轉錄調控因子,進而引發一系列生理、生化反應,形成高效有序的信號調控網絡,從而對脅迫產生應答反應。
作為一種簡單的植物氣體激素,乙烯在植物的生長發育和逆境脅迫應答中發揮重要作用。乙烯參與調控植物的種子萌發、植株生長、開花、果實成熟、單性花的性別決定、器官衰老與脫落等生理過程,此外乙稀還在植物生物與非生物脅迫應答中發揮重要作用。植物體通過轉錄水平和轉錄后水平的協同調控來調節乙烯的生物合成和信號轉導途徑的開閉,協調乙烯信號途徑與其它激素信號途徑的交叉作用,完成各種生命活動和抵御各種逆境脅迫。
乙烯響應因子(ethyleneresponsivefactor,erf)是植物特有的一種轉錄因子,位于乙烯信號通路下游,可調控脅迫相關基因的表達,在植物對環境脅迫的應答方面起到重要的作用。erf不僅能通過結合與病程相關基因啟動子中的一來調節它們的表達從而提高植物的抗病性,還能通過識別和其他非一順式作用元件參與植物的非生物脅迫的應答。
目前,在水稻、小麥、番茄等植物中erf類基因的研究較多,但對桑樹中的erf類基因研究還比較少。
我國是桑樹的起源中心,種質資源極為豐富。經過桑樹種質資源及育種工作者幾十年的努力,現在全國已收集保存有近3000份桑樹種質資源,其中不少資源都具有果用價值、藥用價值、經濟價值等。我們用克隆得到的魯桑品種(morusmulticaulis)育71-1mmerf基因orf序列,通過生物信息學分析其序列特征。同時,利用熒光定量pcr的方法檢測了mmerf基因在干旱、高鹽、低溫、aba脅迫條件下的轉錄水平的變化規律。通過對該基因的研究,使我們從分子水平了解到桑樹mmerf基因在不同脅迫條件下的表達情況。
技術實現要素:
本發明要解決的技術問題是提供一種桑樹乙烯響應因子(mmerf)基因克隆和其在非生物脅迫條件下的表達,可以看桑樹中的乙烯響應因子基因在環境脅迫下的表達調控。
對于桑樹乙烯響應因子基因,本發明采用的技術方案是,其核苷酸序列如seqidno.1所示。
上述桑樹乙烯響應因子基因作為植物非生物脅迫表達標志物中的應用。
作為優選,所述植物為桑樹,其拉丁文學名為morusmulticaulis。
作為優選,所述的非生物脅迫為干旱、高鹽、低溫、aba脅迫。
對于桑樹乙烯響應因子基因編碼的蛋白,本發明采用的技術方案是,其氨基酸序列如seqidn0.2所示。
上述桑樹乙烯響應因子基因編碼的蛋白作為植物非生物脅迫表達標志物中的應用。
作為優選,所述植物為桑樹,其拉丁文學名為morusmulticaulis。
作為優選,所述的非生物脅迫為干旱、高鹽、低溫、aba脅迫。
本發明的有益效果是:
本發明完善桑樹基因組同時探究桑樹乙烯響應因子(mmerf)基因在非生物脅迫條件的表達分析,桑樹mmerf的克隆將為進一步通過轉基因技術培育抗性桑樹新品種奠定基礎,可有效彌補常規育種技術的不足,從而加速桑樹抗性新品種的選育進程。
附圖說明
下面結合附圖和具體實施方式對本發明作進一步詳細的說明。
圖1為熒光定量rt-pcr檢測mmerf基因在高鹽脅迫下桑苗嫩葉中的相對表達量圖;ck:對照;1d:高鹽處理1天;2d:高鹽處理2天;4d:高鹽處理4天;7d:高鹽處理7天;10d:高鹽處理10天。
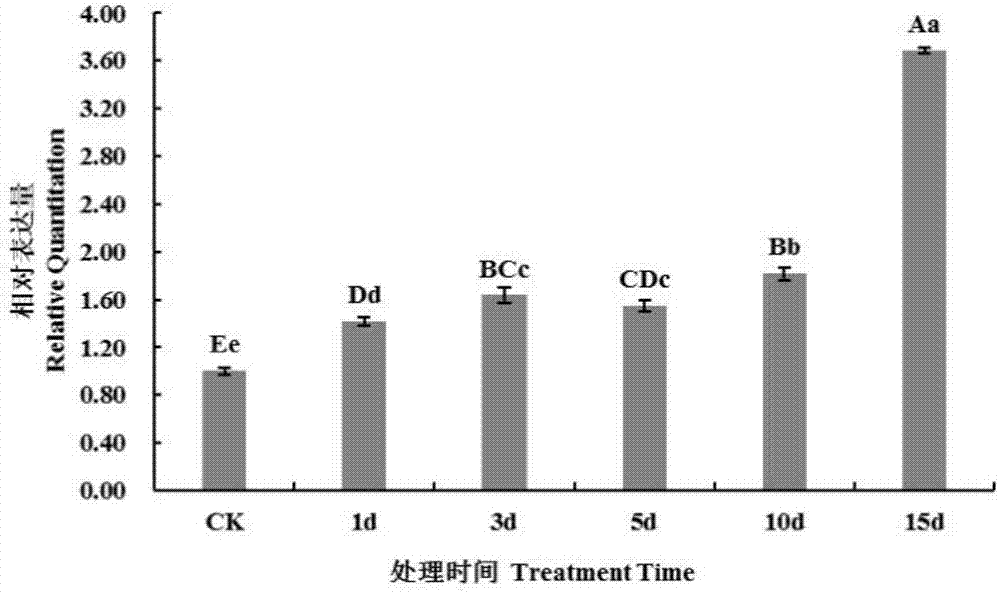
圖2為熒光定量rt-pcr檢測mmerf基因在干旱脅迫下桑苗嫩葉中的相對表達量圖;ck:對照;1d:干旱處理1天;3d:干旱處理3天;5d:干旱處理5天;10d:干旱處理10天;15d:干旱處理15天。
圖3為熒光定量rt-pcr檢測mmerf基因在4℃低溫脅迫下桑苗嫩葉中的相對表達量圖;ck:對照;12h:干旱處理12小時;1d:干旱處理1天;3d:干旱處理3天;6d:干旱處理6天;10d:干旱處理10天;rn2d:常溫復性2天;rn5d:常溫復性5天。
圖4為熒光定量rt-pcr檢測mmerf基因在aba脅迫下桑苗嫩葉中的相對表達量圖;ck:對照;2h:aba處理2小時;6h:aba處理6小時;12h:aba處理12小時;1d:aba處理1天;3d:aba處理3天;5d:aba處理5天。
具體實施方式
實施例1桑樹乙烯響應因子mmerf基因的克隆
1.1設計引物
供試桑樹品種為保存于中國農業科學院蠶業研究所國家種質鎮江桑樹圃的育71-1。利用已獲得的包含mmerf基因cdna片段的ests序列,設計引物克隆,并加以驗證,引物序列如下:
mmmmerf-f:5′-aggaaaacaaatttctcgtca-3′;
mmmmerf-r:5′-aacgcaagttttcaaggtc-3′
根據已獲得的基因orf序列設計進行熒光定量分析的引物,引物序列如下:
qmmerf-f:5′-ttccagggtttcaaggatgac-3′
qmmerf-r:5′-gttgttgagccacgagagaa-3′
根據ncbi上查找的在桑樹中穩定表達的β-actin基因(genebank登錄號:dq785808)序列設計內參基因的上下游引物,上游引物序列β-actin-f為5′-agcaactgggatgacatggaga-3′,下游引物序列β-actin-r為5′-cgaccactggcgtaaaggga-3′。
1.2桑樹乙烯響應因子mmerf基因的克隆
使用takara試劑盒提取桑樹嫩葉rna,提取步驟按照試劑盒說明書進行,電泳檢測rna條帶完整;并按照takara反轉錄試劑盒說明書將rna反轉錄成cdna。以合成的cdna第1鏈為模板,利用mmerf-f/r為引物進行pcr擴增,其中,pcr酶使用takara公司生產的extaq酶,pcr反應程序為:94℃預變性5min;94℃變性30s,55℃退火35s,72℃延伸40s,37個循環;72℃復性5min;4℃保存。電泳檢測目的片段大小,使用takara公司agarosege1dnapurificationkit試劑盒純化回收pcr產物,得到桑樹mmerf基因片段,純化方法參照試劑盒說明書。將目的片段用dna連接酶與pmdtm18-t載體連接。連接產物轉化到宿主菌e.colitop10菌株感受態細胞,涂布于在含有amp的lb固體平板上。挑取單菌落培養后送樣,由生工生物工程(上海)有限公司完成測序,測序結果進行blast分析,獲得1500bp的包含完整orf的mmerf基因序列(genbankaccessionnumber:mf188168),其中,包括140bp的5′-utr,193bp的3′-utr和1167bp的完整orf。
實施例2桑樹乙烯響應因子mmerf基因的生物信息學分析
利用在線分析工具psort和smart(http://smart.embl-heidelberg.de/),根據已報道的文獻對目的基因進行核定位序列的預測及蛋白質結構域的分析,桑樹乙烯響應因子mmerf蛋白有一個由64個氨基酸殘基組成的ap2/erf結構域,位于蛋白的中部,其中,第14位和第19位氨基酸殘基分別為丙氨酸(a)和天冬氨酸(d),說明它為典型的erf轉錄因子。
利用dnastar軟件和在線分析軟件expasy(http:∥prosite.expasy.org/prosite.html/)對桑樹乙烯響應因子mmerf基因進行較全面的分析,獲得該基因所編碼的氨基酸序列seqidno.2,基因編碼桑樹乙烯響應因子mmerf蛋白質的等電點為5.02,分子質量為43.30kda。
實施例3桑樹乙烯響應因子mmerf基因在各種脅迫條件下的表達模式
在供試桑苗的新梢生長至約20cm長時,對桑苗分別進行干旱、高鹽、低溫、aba脅迫處理。在整個脅迫誘導處理過程中,所有試驗用苗的溫度,光照強度,光周期以及濕度等非生物條件均保持不變。當桑苗在各種脅迫處理下表現出明顯受害癥狀時,分別取對照和各種脅迫條件下的桑樹幼葉,用鋁箔紙包裹后迅速投入液氮中,放于-80℃的冰箱中保存。采用熒光定量pcr的方法,以桑樹穩定表達的肌動蛋白基因(β-actin)作為內參,測定桑樹在脅迫條件下目的基因的表達水平。所有脅迫實驗均為3個生物學重復3個技術重復。
高鹽脅迫:供試桑苗用濃度為300mm的nacl溶液進行鹽脅迫誘導處理。試驗桑苗和對照桑苗放置于同一個光照培養箱,溫度25℃,每天16小時光照,8小時黑暗。分別于1天、2天、4天、7天和10天時取鹽脅迫條件下的桑樹幼葉(第1-3葉位)作為實驗材料。
干旱脅迫:供試桑苗放置于溫度25℃光照培養箱,一直不澆水,每天16小時光照,8小時黑暗。分別于1天、3天、5天、10天和15天取桑樹幼葉(第1-3葉位)作為實驗材料。
低溫脅迫:供試桑苗放置于溫度4℃光照培養箱,每天16小時光照,8小時黑暗。分別于12小時、1天、3天、6天和10天取桑樹幼葉(第1-3葉位)作為實驗材料,后放置于溫度25℃光照培養箱進行常溫復性,每天16小時光照,8小時黑暗,復性2天和5天取桑樹幼葉(第1-3葉位)作為實驗材料。
aba脅迫:供試桑苗用濃度為0.1mol/l的aba溶液進行aba脅迫誘導處理,放置于溫度25℃光照培養箱,每天16小時光照,8小時黑暗。分別于2小時、6小時、12小時、1天、3天和5天取桑樹幼葉(第1-3葉位)作為實驗材料。
不同非生物脅迫下桑樹乙烯響應因子mmerf基因相對表達量變化顯示:高鹽脅迫條件下,mmerf基因相對表達量均比正常生長條件下低,總體的趨勢是先緩慢下降,在處理4天時降到最小值(對照的0.52倍),然后再緩慢上升,最后到第10天上升到對照的0.85倍(圖1)。干旱脅迫條件下,mmerf基因相對表達量整體趨勢表現上調,隨著干旱處理時間的延長,mmerf基因相對表達量逐漸升高,在第15天時達到最大值,為對照的3.69倍(圖2)。低溫脅迫條件下,mmerf基因的相對表達量整體趨勢表現明顯上調,持續處理1天后上調到較高值(對照的2.53倍),然后緩慢下降后急速上升到這幾個階段的最大值(對照的3.89倍),經常溫復性2天,表達量回落,繼續復性5天后,表達量恢復到接近對照值(圖3)。aba脅迫條件下,mmerf基因相對表達量整體趨勢表現下調,隨著aba脅迫處理時間的延長,mmerf基因相對表達量先是逐漸下降,在第12小時降到最小值,為對照的0.33倍,后緩慢升高,到第5天保持在對照的0.78倍左右(圖4)。mmerf基因在各種脅迫條件下都有明顯的波動狀態,這表明mmerf可能發揮其功能,在非生物脅迫條件下,誘發mmerf基因的表達,使植物積極響應外界脅迫。通過熒光定量pcr分析,利用mmerf基因在干旱脅迫、高鹽脅迫、低溫脅迫和aba脅迫條件下總體表達的升降情況、在不同的脅迫時間內相對基因表達量指標,以及mmerf基因在植物中的保守性,可以以此標識植物是否處于高鹽、低溫、干旱或aba脅迫等生長環境。
以上所述的本發明實施方式,并不構成對本發明保護范圍的限定。任何在本發明的精神和原則之內所作的修改、等同替換和改進等,均應包含在本發明的權利要求保護范圍之內。
sequencelisting
<110>安徽省農業科學院蠶桑研究所
<120>桑樹乙烯響應因子mmerf及其應用
<130>no
<160>2
<170>patentinversion3.3
<210>1
<211>1500
<212>dna
<213>mmerf
<400>1
aggaaaacaaatttctcgtcaacaaaaaaaaaaaaaaaaaaaaaaaaaaactttcttctc60
tgaaaaatctcaaaccttttttcttgattctctgcaagaggtgttttggatccgggttgt120
ttgttgtgcagtgaaggagaatgtgtggaggtgctattatctccgacttcatctcgacgc180
cgcggtccacgcgtctgaccgcggactacctctggccggatctgaagaaatccggctccg240
gcaagcgcttctgcaagcctgtgagatcggtgatcgtcgacatcgacgatgacttcgagg300
ctgatttccagggtttcaaggatgactccgacgtcgacaacgacgacgacgttatagatg360
tcaagcctttcgctttctctgctcgcaagcccaccttctctcgtggctcaacaactgtga420
aatataccgaatctgatgggcaagctgagaaatctgcaaagagaaagagaaagaaccagt480
ataggggaattcggcagcgcccatggggtaagtgggctgctgagatccgcgaccctagga540
aaggtgtccgggtttggctcggaactttcaacactgctgaagaagctgcaagagcttatg600
atgctgaggcacgacgaatccgtggtaagaaagccaaggtgaatttcccagatgaaactc660
cgcgtgctttgcctaaacatcctgttaaggaaagtcctaagagatcactgcccaaggaaa720
attcgaactccaccgagtcgaatttgaacaatcaaagtttcaattttgtgaataactctg780
atctggactactacaatgctatgggttttctggaagagaaaccgctgactaatcaatatg840
agcatgtggagactctccctgctaaagcggatgttggactcaaatccaatgctccagctg900
ctactactccgctgtatttcagttctgatcagggtagcaattcatttgagtgttccgact960
ttgggttgggagaacatggctcaaagactccagaaatttcttctgttttctcagccactt1020
cagaaaatgatgactcgctatttcttgaggatactaacccaacaaagaagctgaagtctg1080
actcagagaatgcggtgcttcctgaagaaaatcatgcaaagactctgtcagaggagcttt1140
cggctttcgagtctcagatgaagttctttcagatgccatatcttgagggaagctgggatg1200
cgttattggacaccttcctcgctggcgattcaactcaggatggtggaaactctatcaatc1260
tttggagttttgatgacttctccaccgtgtctggcgaagccttctaagccaattccccag1320
ctttcctaattaatgtaaataaggctacatgaattgtttatctgcataaggattgcacaa1380
acagatgaacatgcacaagccttgtatggagagtgattcgaggtagctaatatcggtgta1440
tggcgagtatatagatattaggttttacttcttaccttgaagaccttgaaaacttgcgtt1500
<210>2
<211>388
<212>蛋白質1
<213>mmerf蛋白
<400>1
mcggaiisdfistprstrltadylwpdlkksgsgkrfckpvrsvivdidddfeadfqgfk60
ddsdvdndddvidvkpfafsarkptfsrgsttvkytesdgqaeksakrkrknqyrgirqr120
pwgkwaaeirdprkgvrvwlgtfntaeeaaraydaearrirgkkakvnfpdetpralpkh180
pvkespkrslpkensnstesnlnnqsfnfvnnsdldyynamgfleekpltnqyehvetlp240
akadvglksnapaattplyfssdqgsnsfecsdfglgehgsktpeissvfsatsenddsl300
fledtnptkklksdsenavlpeenhaktlseelsafesqmkffqmpylegswdalldtfl360
agdstqdggnsinlwsfddfstvsgeaf388
- 還沒有人留言評論。精彩留言會獲得點贊!