一種表達Streptomycessp.FA1來源木聚糖酶的畢氏酵母重組菌的制作方法
本發明涉及一種表達streptomycessp.fa1來源木聚糖酶的畢氏酵母重組菌,屬于基因工程領域。
背景技術:
木聚糖酶屬于糖苷水解酶[ec3.2.1.x],能應用在多個領域中。木聚糖酶和其他半纖維素酶一起添加在飼料中利于增加飼料營養分解;在制漿和造紙工業領域,木聚糖酶可作為預漂白劑處理紙漿,從而降低紙漿漂白過程中化學試劑使用量,減少對環境的污染;在食品領域可以用于面制品烘焙,能夠改善面制品的品質;木聚糖酶還用于制備低聚木糖。麻類植物的纖維素含量豐富,是紡織行業中纖維原材料,但是麻類植物中含有半纖維素、木質素、果膠等雜質不利于紡織,木聚糖酶可以進行脫膠以去除這些膠質,因而在紡織工業中木聚糖酶也有相當的應用,可減少化學品對環境的污染。迄今為止,國內外已報道了400多種木聚糖酶基因,大多數木聚糖酶在野生菌中表達量僅在10-20iu·ml-1。為了進一步推動木聚糖酶在工業生產中的應用,利用分子生物學方法將木聚糖酶基因進行異源表達以提高木聚糖酶的表達水平,是目前普遍采用的木聚糖酶制備方式。
畢氏酵母表達系統主要包括誘導型表達系統和組成型表達系統。誘導型表達系統中使用乙醇氧化酶為啟動子(paox1)來誘導表達外源蛋白,期間需要進行初始碳源和甲醇之間的轉換,導致存在發酵周期長、操作不方便、需要掌握好碳源的轉換時間、甲醇含量難以監測的缺點;此外甲醇屬于易揮發、易燃易爆物質,具有一定的生產危險性,不方便大量儲存;甲醇還屬于劇毒物質,不適用于食品方面的應用,因而制約了paox1表達載體在食品領域的大規模應用。組成型表達系統是最近幾年廣泛使用的畢氏酵母表達系統,其中三磷酸甘油醛脫氫酶啟動子(pgap)是較為常用的同生長相偶聯的組成型表達系統啟動子,可以利用甘油或者葡萄糖等碳源生長并高效表達外源基因,具有不需要更換碳源、操作便捷的優點。尤其是該啟動子利用無毒害性碳源,使pgap質粒構建的組成型表達系統對于表達食品行業用酶制劑具有更重要意義。
外源基因可以整合進宿主基因組進行表達,也能以游離質粒的形式進行表達。整合型表達具有遺傳穩定的優點,而游離型表達具有拷貝數高的優點。目前在畢氏酵母表達系統中,木聚糖酶基因均是構建整合型重組菌株進行外源基因重組表達,尚未見到采用游離型表達質粒來進行表達。
技術實現要素:
為解決上述技術問題,本發明提供了一種穩定的、可以大量分泌木聚糖酶的游離表達型畢氏酵母重組菌及其構建方法,工業應用前景廣闊。
本發明的第一個目的是提供一種表達木聚糖酶的畢氏酵母重組菌,所述畢氏酵母重組菌是將streptomycessp.fa1來源的木聚糖酶基因,連接到含有自主復制序列的游離表達質粒上,轉化至畢氏酵母中得到的畢氏酵母重組菌。
在本發明的一種實施方式中,所述木聚糖酶基因氨基酸序列如seqidno:1所示。
在本發明的一種實施方式中,所述自主復制序列的核苷酸序列如seqidno:2所示。
在本發明的一種實施方式中,所述游離表達載體為通過將核苷酸序列如seqidno:2的pars復制子連接到pgapzαa質粒上得到pgapzαa-pars游離表達載體。
在本發明的一種實施方式中,所述畢氏酵母為畢氏酵母km71、gs115或smd168。
本發明第二個目的是提供一種所述畢氏酵母重組菌的構建方法,所述方法包括如下步驟:
(1)獲得氨基酸序列如seqidno:1所示的木聚糖酶基因;
(2)獲得序列如seqidno:2所示的自主復制序列pars基因;
(3)將步驟(2)所得pars基因與質粒pgapzαa相連,得到表達載體pgapzαa-pars;
(4)將步驟(1)中的木聚糖酶基因與步驟(3)中的表達載體pgapzαa-pars相連,得到重組載體pgapzαa-pars-xyna;
(5)將重組載體pgapzαa-pars-xyna轉化到畢氏酵母中,得到所述畢氏酵母重組菌。
本發明的第三個目的是提供所述的畢氏酵母重組菌生產木聚糖酶的方法,所述方法包括以下步驟:
(1)以權利要求1所述畢氏酵母重組菌為生產菌株,活化后,30℃、200rpm條件下培養24h,得一級種子發酵液;
(2)以2.5%接種量接種一級種子發酵液至種子培養基中,30℃、200rpm條件下培養24h,得二級種子發酵液;
(3)以10%的接種量將二級種子發酵液接種至發酵培養基中30℃、200rpm進行發酵;
(4)待do值迅速上升時,進行補料,od600=100時停止補料,饑餓至第二次出現do值迅速上升時繼續補料。
在本發明的一種實施方式中,所述種子培養基為ypd培養基;所述發酵培養基為bsm培養基;其中bsm培養基包括如下成分:85%磷酸26.7ml/l,caso40.93g/l,k2so418.2g/l,mgso4·7h2o14.9g/l,koh4.13g/l,甘油40.0g/l,ptm14.35ml/l,上述成分均溶于水中形成所述bsm培養基。
在本發明的一種實施方式中,所述方法中補料配方為50%甘油、1.25%ptm1;甘油和ptm1溶于水中形成所述補料。
本發明的第四個目的是提供所述畢氏酵母重組菌在飼料、食品、紡織或化工領域的應用。
本發明的有益效果
本發明所述重組畢氏酵母工程菌屬于帶自我復制的游離型載體重組畢氏酵母工程菌。此工程菌相對通用的整合型的重組畢氏酵母工程菌顯著提升了分泌生產木聚糖酶的能力。此外發酵原料來源廣泛,生產成本較低。
本發明將streptomycessp.fa1來源木聚糖酶(xyna)基因整合到游離含pgap啟動子的質粒中再轉入畢氏酵母,使其分泌木聚糖酶。同整合型表達菌株相比,采用游離載體表達木聚糖酶產量提高了50%,同時表達的酶的比活力提高了90%,從而顯著降低了工業化生產木聚糖酶的生產成本。
附圖說明
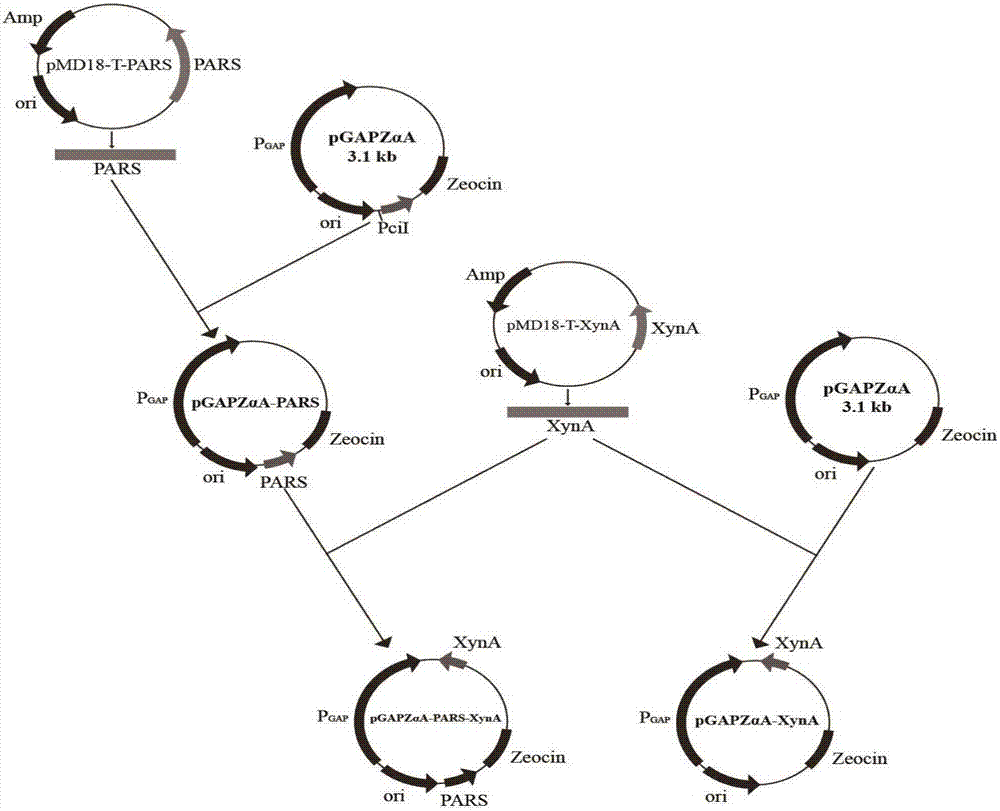
圖1重組質粒pgapzαa-pars-xyna的構建過程圖;
圖2pgapzαa-pars-xyna重組質粒的雙酶切驗證圖;
圖3pgapzαa-pars-xyna重組菌3.6l發酵sds-page電泳圖;
圖4pgapzαa-pars-xyna重組菌3.6l發酵酶活對比圖。
具體實施方式
將0.5g的木聚糖溶于100ml,50mmol/l,ph5.5的磷酸緩沖液充分混勻,取1ml底物,在55℃預熱10min,加入1ml的酶液,反應10min后加入3mldns,煮沸10min迅速冷卻,加蒸餾水定容至20ml,540nm下測吸光度(以滅活的酶液為催化劑同樣操作作為空白對照)。
在上述條件下,將每分鐘水解木聚糖糖生成1μmol的木糖所需酶量定義為木聚糖酶的一個單位的酶活力(u)。
實施例1:表達載體pgapzαa-pars的構建
利用全基因合成技術合成核苷酸序列如seqidno.2所示的自主復制序列pars;將回收得到的pars基因片段,用infusion酶與質粒pgapzαa相連,構建得到表達載體pgapzαa-pars。
實施例2:游離型pgapzαa-pars-xyna重組菌的構建
重組質粒pgapzαa-pars-xyna的構建:
以質粒pmd18-t-xyna(axylanasefromstreptomycessp.fa1:heterologousexpression,characterization,anditsapplicationinchinesesteamedbread,yangxu,journalofindustrialmicrobiology&biotechnology,may2016,volume43,issue5,pp663–670)為模板,設計序列如seqidno.3所示的正向引物:ccggaattcatggccgagaacaccctt,序列如seqidno.4所示的反向引物:atttgcggccgctcaggtgcgggtccagcgtt。通過聚合酶鏈式反應獲得具有noti和ecori酶切位點的xyna片段。
pcr反應體系(50μl):
pcr程序:94℃,4min(預變性);98℃,10s(變性);60℃,5s(退火);72℃,90s(延伸);設置30個循環;72℃,10min(保溫)后設置保藏溫度為4℃。將pcr產物進行膠回收、酶切回收目的基因將其與表達載體pgapzαa-pars進行雙酶切,并于16℃連接過夜,轉化e.colijm109,涂布含zecoin抗性的lb平板,37℃培養8-10h,挑取轉化子,提取重組質粒并雙酶切驗證,然后對驗證正確的重組質粒測定dna序列,陽性克隆子即pgapzαa-pars-xyna。見圖1和圖2。
重組質粒pgapzαa-pars-xyna的轉化:
(1)從乙醇中取出電轉杯,置于超凈臺中吹干,并置于冰中預冷備用;
(2)在冰上進行以下操作:將重組游離質粒pgapzαa-pars-xyna預冷,吸取5μl快速與80μl酵母感受態中混勻,轉移到0.2cm經過紫外照射的的電轉杯,冰浴5min,電擊條件為:電壓1500v,時間控制4-10ms;
(3)迅速向電擊杯中加入1ml1mol·l-1預冷的山梨醇,輕輕吹打均勻,然后將其轉移至冰冷的1.5ml無菌ep管中,30℃、50r·min-1培養1-2h;
(4)離心菌體,重新懸浮均勻后吸取200μl-300μl涂布上還有zecoin抗性的ypd平板上,于30℃恒溫條件下培養,至長出單菌落,即為含有游離質粒的重組單菌落。
(5)使用無菌牙簽挑取ypd平板上的菌落,接種到ypd培養基中。
(6)從平板上挑取陽性轉化子單酵母菌落轉接于裝有10mlypd培養基的50ml三角瓶中培養24h后轉入裝有50mlypd培養基的500ml三角瓶培養表達3~4天,培養物經離心后得到含重木聚糖酶的上清,利用dns法對其進行酶活檢測。
實施例3:整合型pgapzαa-xyna重組菌的構建
重組質粒pgapzαa-xyna構建:
以質粒pmd18-t-xyna為模板,設計序列如seqidno.3所示的正向引物:ccggaattcatggccgagaacaccctt,序列如seqidno.4所示的反向引物:atttgcggccgctcaggtgcgggtccagcgtt。通過聚合酶鏈式反應獲得具有noti和ecori酶切位點的xyna片段。
pcr反應體系(50μl):
pcr程序:94℃,4min(預變性);98℃,10s(變性);60℃,5s(退火);72℃,90s(延伸);設置30個循環;72℃,10min(保溫)后設置保藏溫度為4℃。將pcr產物進行膠回收、酶切回收目的基因將其與表達載體pgapzαa進行雙酶切,并于16℃連接過夜,轉化e.colijm109,涂布含zecoin抗性的lb平板,37℃培養8-10h,挑取轉化子,提取重組質粒并雙酶切驗證,然后對驗證正確的重組質粒測定dna序列,陽性克隆子即pgapzαa-xyna。
重組質粒pgapzαa-xyna的轉化:
重組質粒經saci內切酶線性化,酶切體系如下:
混勻上述溶液,37℃酶切2h,采用瓊脂糖凝膠電泳驗證酶切得到產物。
(1)從乙醇中取出電轉杯,置于超凈臺中吹干,并置于冰中預冷備用
(2)在冰上進行以下操作:將線性化后回收的pgapzαa-xyna預冷,吸取5μl快速與80μl酵母感受態中混勻,轉移到0.2cm經過紫外照射的的電轉杯,冰浴5min,電擊條件為:電壓1500v,時間控制4-10ms;
(3)迅速向電擊杯中加入1ml1mol·l-1預冷的山梨醇,輕輕吹打均勻,然后將其轉移至冰冷的1.5ml無菌ep管中,30℃、50r·min-1培養1-2h;
(4)離心菌體,重新懸浮均勻后吸取200μl-300μl涂布上還有zecoin抗性的ypd平板上,于30℃恒溫條件下培養,至長出單菌落,即為整合到基因組的重組單菌落。
(5)使用無菌牙簽挑取ypd平板上的菌落,接種到ypd培養基中。
(6)從平板上挑取陽性轉化子單酵母菌落轉接于裝有10mlypd培養基的50ml三角瓶中培養24h后轉入裝有50mlypd培養基的500ml三角瓶培養表達3~4天,培養物經離心后得到含重木聚糖酶的上清,利用dns法對其進行酶活檢測。
實施例4:pgapzαa-pars-xyna重組菌與pgapzαa-xyna重組菌3.6l發酵罐發酵
重組畢氏酵母工程菌發酵罐發酵生產木聚糖酶的方法在3.6linfors罐進行,過程分為三階段進行:
第一階段:接種實施例3或實施例5中的陽性轉化子單菌落至裝有10mlypd培養基的50ml三角瓶中,200rpm,30℃振蕩培養24h,作為一級種子發酵液;
第二階段:取所述一級種子發酵液2.5ml接種于裝有100ml發酵培養基的500ml三角瓶中,200rpm,30℃振蕩培養24h,作為二級種子發酵液;
第三階段:將100ml的種子發酵液接種到裝有900mlbsm培養基中的3.6l發酵罐中,200rpm,30℃的條件下培養,全程用25%氨水進行ph值的調節,保持在5.0;所述bsm培養基包括如下成分:85%磷酸26.7ml/l,caso40.93g/l,k2so418.2g/l,mgso4·7h2o14.9g/l,koh4.13g/l,甘油40.0g/l,ptm14.35ml/l上述成分均溶于水中形成所述bsm培養基;
待碳源耗盡,即do值迅速上升時,進行補料,待od600=100時,停止補料,饑餓至第二次出現do值迅速上升時,繼續補料,補料配方為50%甘油;1.25%ptm1;甘油和ptm1溶于水中形成所述補料。不同時間段取樣。
實施例5:木聚糖酶酶活測定
分別取實施例6中pgapzαa-pars-xyna重組菌與pgapzαa-xyna重組菌產的酶進行酶活測定,實驗結果見圖4。結果表明,整合型pgapzαa-xyna重組菌獲得的木聚糖酶的最大酶活力235u/ml,蛋白含量2.7mg/ml。游離型pgapzαa-pars-xyna重組菌獲得的木聚糖酶的最大酶活力為350u/ml,蛋白含量2.1mg/ml,同整合型pgapzαa-xyna重組菌相比,采用游離載體表達木聚糖酶產量提高了50%,同時表達的酶的比活力提高了90%,從而顯著降低了工業化生產木聚糖酶的生產成本。此外,pgapzαa-pars-xyna重組菌所產酶的蛋白電泳結果顯示在43kda處有一條與理論分子量一致的條帶(結果如圖3所示)。
雖然本發明已以較佳實施例公開如上,但其并非用以限定本發明,任何熟悉此技術的人,在不脫離本發明的精神和范圍內,都可做各種的改動和修飾,因此本發明的保護范圍應該以權利要求書所界定的為準。
序列表
<110>江南大學
<120>一種表達streptomycessp.fa1來源木聚糖酶的畢氏酵母重組菌
<160>4
<170>patentinversion3.3
<210>1
<211>436
<212>prt
<213>streptomycessp.fa1
<400>1
metalagluasnthrleuglyalaalaalaalaglnserglyargtyr
151015
pheglyvalalailealaserglylysleuglyaspserthrtyrthr
202530
serilealaasnargglupheasnservalthralagluasnglumet
354045
lysileaspalathrgluproasnargglyglnpheasnpheserser
505560
alaaspargvaltyrasntrpalavalglnasnglylysglnvalarg
65707580
glyhisthrleualatrphisserglnglnproglytrpmetglnser
859095
leuserglyserserleuargglnalametileasphisileasngly
100105110
valmetalahistyrlysglylysilealaglntrpaspvalvalasn
115120125
glualaphealagluglyserserglyalaargargaspserasnleu
130135140
glnargthrglyasnasptrpilegluvalalapheargthralaarg
145150155160
alaalaaspproseralalysleucystyrasnasptyrasnvalglu
165170175
asntrpthrtrpalalysthrglnalamettyrasnmetvallysasp
180185190
phelysserargglyvalproileaspcysvalglypheglnserhis
195200205
pheasnserglyserprotyrasnserasnpheargthrthrleugln
210215220
asnphealaalaleuglyvalaspvalalavalthrgluleuaspile
225230235240
glnglyalaserserserthrtyralaalavalvalasnaspcysleu
245250255
alavalserargcysleuglyvalthrvaltrpglyvalargaspser
260265270
aspsertrpargalaseraspthrproleuleupheasnasnaspgly
275280285
serlyslysalaalatyrseralavalleuasnalaleuasnglygly
290295300
thrthrthrproproprothrglyaspglyglyglnilelysglyval
305310315320
alaserglyargcysleuaspvalproasnalaserthrthraspgly
325330335
thrglnileglnleutyraspcyshisserasnserasnglnglntrp
340345350
alavalthraspserglygluileargvaltyrglyasplyscysleu
355360365
aspalaalaglythrglyasnglyalaprovalglniletyrsercys
370375380
trpglyglyaspasnglnlystrpargleuasnseraspglyserile
385390395400
valglyvalglnserglyargcysleuaspalaalaglythrglyasn
405410415
glyalaargileglnleutyrsercysserglyglyserasnglnarg
420425430
trpthrargthr
435
<210>2
<211>164
<212>dna
<213>人工合成
<400>2
tcgagataagctgggggaacattcgcgaaaatgaaacaagtcggctgttatagtatattt60
attataatattgaaagatctcaaaagactacttatttttgaatgaaccaagtatgaaatc120
aacctatttggggttgaccaaaataagtaaatattaattgtcga164
<210>3
<211>27
<212>dna
<213>人工合成
<400>3
ccggaattcatggccgagaacaccctt27
<210>4
<211>32
<212>dna
<213>人工合成
<400>4
atttgcggccgctcaggtgcgggtccagcgtt32
- 還沒有人留言評論。精彩留言會獲得點贊!