ZmRCI2?8基因及其在促進非生物脅迫條件下植物發芽和側根生長中的應用的制作方法
本發明屬于基因工程技術和生理遺傳領域,具體涉及一種可響應多種非生物脅迫的蛋白的基因。
背景技術:
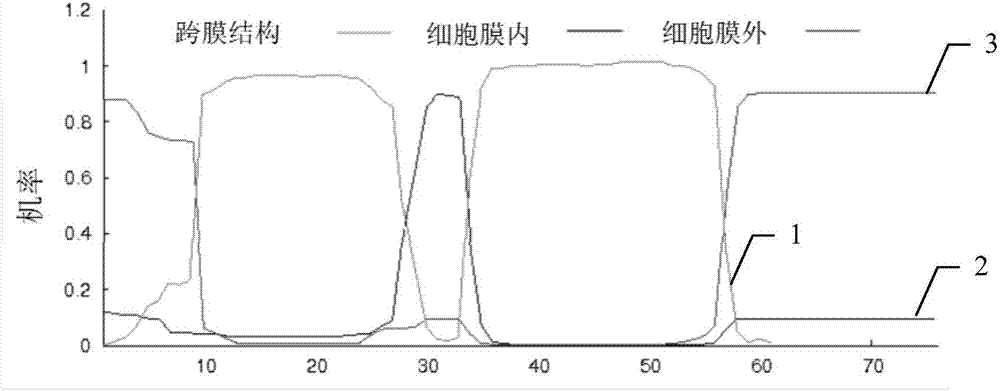
:磷是多種有機化合物的重要組分,包括核酸、磷脂、atp等。磷參與植物體內碳水化合物代謝、氮素代謝及脂肪代謝等生理過程,還調控酶促反應及細胞信號的轉導,并且能夠通過蛋白修飾調控多種重要的生理和生物學過程。同時,磷可以促進根系伸長,降低葉片蒸騰強度,提高植物忍受干旱傷害的能力,或維持和調控作物體內新陳代謝過程,提高植物抗寒性。磷是大量元素,但植物能從土壤中吸收的有效磷卻很少,世界上至少三分之一可耕作土壤中的磷不能被植物吸收,因此磷被認為是影響植物生長和作物產量的主要限制因素。另一方面,而且,磷肥也不是施用越多越好。如果磷肥施用過度,還會存在如下問題:(1)過多磷素營養促使作物呼吸作用過旺,造成繁殖器官提前發育,引起作物過早成熟,籽粒小,產量低。(2)誘發土壤缺鋅。土壤里的鋅與過量的磷作用,產生磷酸鋅沉淀,或者過量的磷導致土壤堿化,使鋅的有效性降低。(3)使作物缺硅。據研究,過量磷肥會造成土壤中的硅被固定,不能吸收,引起作物缺硅,導致莖稈纖細,倒伏及抗病能力差等缺硅癥狀,特別是對喜硅的禾本科作物例如玉米的影響更大。(4)使作物得磷缺鉬,表現出“缺鉬癥”。(5)造成土壤中有害元素積累。磷肥包含鎘、鉛、氟等有害元素,施用磷肥會引起土壤中鎘等的增加,其年增長量甚至達到0.15%,且鎘有效性高,易為作物吸收,給人畜造成危害。(6)造成土壤理化性質惡化,造成土壤酸化(過磷酸鈣)或堿性加重(鈣鎂磷肥)以及理化性質惡化。植物需要從土壤中吸收磷來維持生長發育。在缺磷條件下,植物可以通過光合作用、碳代謝等代謝調節來降低體內磷的消耗,從而提高磷的利用效率。為了最大程度地利用磷源,在缺磷條件下,可以利用磷酸酶、核酸酶等活性物質將植物體內的磷從衰老的組織器官轉移到新生組織中。植物對低磷脅迫響應的適應性變化過程是一系列基因時空特異表達的結果。低磷脅迫后,水稻、玉米、白羽扇豆和擬南芥等植物中的大量基因差異性表達,這些差異表達基因涉及植物多種生理生化途經(wasakij等.2006.transcriptomicanalysisindicatesputativemetabolicchangescausedbymanipulationofphosphorusavailabilityinriceleaves(轉錄物組學分析表明控制水稻葉子中的磷利用度導致推測的代謝變化).journalofexperimentalbotany57:2049-2059;morcuender等.2007.genome-widereprogrammingofmetabolismandregulatorynetworksofarabidopsisinresponsetophosphorus(擬南芥中對磷響應的代謝和調控網絡的基因組范圍內的重新編程).plantcellandenvironment30:85-112;hernandezg等.2007.phosphorusstressincommonbean:roottranscriptandmetabolicresponses(菜豆中的磷脅迫:根轉錄和代謝應答).plantphysiology144:752-767.)。低磷響應基因可根據其缺磷時間長短分為兩類,一類是早期快速響應基因,這類基因對缺磷反應敏感,缺磷處理幾小時內基因即表達(hammondjp等.2003.changesingeneexpressioninarabidopsisshootsduringphosphatestarvationandthepotentialfordeveloping(擬南芥嫩條在磷饑餓過程中的基因表達變化和發育潛力).plantphysiology132:578-596.),如轉錄因子atphr1(rubiov等.2001.aconservedmybtranscriptionfactorinvolvedinphosphatestarvationsignalingbothinvascularplantsandinunicellularalgae(在微管植物和單細胞藻類生物中參與磷酸鹽饑餓信號轉導的保守myb轉錄因子).genesdevelopment15:2122-33.),也包括一些啟動子區域可能有pho和tata-box元件的非特異性響應基因(邢國芳,郭平毅.2013.植物響應低磷脅迫的功能基因組研究進展.生物技術通報7:1-6.);另一類是晚期低磷脅迫響應基因,它們大多數為缺磷特異性誘導基因,長時間受低磷脅迫,與植物形態、生理和代謝反應密切相關(hammondjp,broadleymr,whitepj.2004.geneticresponsestophosphorusdeficiency(磷缺乏的基因響應).annalsofbotany(london)94:323-332.),如酸性磷酸酶、磷酸鹽轉運蛋白等(liuc等.1998.tomatophosphatetransportergenesaredifferentiallyregulatedinplanttissuesbyphosphorus(磷在植物組織中的對番茄磷酸鹽轉運子基因的差異調控).plantphysiology116:91-99.)。rci2(rare-cold-inducible)是一類普遍存在于原核生物與真核生物中的高度保守的小分子疏水多肽。rci2/pmp3s家族基因存在于原核生物和真核生物基因組(medinaj等.2007.phylogeneticandfunctionalanalysisofarabidopsisrci2genes(擬南芥rci2基因的系統發生和功能分析).journalofexperimentalbotany58:4333-4346.)。pmp3家族蛋白是以含有55個氨基酸殘基的酵母pmp3p蛋白命名的。植物rci2/pmp3s的大小(50-80個氨基酸殘基)、序列和結構均與pmp3p相似,包括兩個高度疏水性區域和完整的膜蛋白結構,但rci2/pmp3s在細胞離子穩態活性的機制尚不清楚(navarrec和goffeaua.2000.membranehyperpolarizationandsaltsensitivityinducedbydeletionofpmp3,ahighlyconservedsmallproteinofyeastplasmamembrane(一種酵母細胞質膜的高度保守的小蛋白pmp3缺失誘導的膜超極化和鹽敏感性).embojournal19:2515-2524.)。許多植物pmp3基因根據擬南芥atri2a/b命名為罕見低溫誘導基因(rci2或rci2樣(rci2-like))(capelj等.1997.twohomologouslow-temperature-induciblegenesfromarabidopsisencodehighlyhydrophobicproteins(來自擬南芥的兩種同源低溫可誘導的基因編碼高度疏水的蛋白).plantphysiology115:569-576.)。rci2基因參與響應多種非生物脅迫。根據rci基因的遺傳特性和超表達研究發現,多數rci2基因可能參與調節脅迫耐受性、細胞離子平衡、穩定膜結構等。然而,rci2基因受脅迫表達有時候上調,有時候表達下調。丹參smpmp3-1/-2在苗期受鹽脅迫和aba處理均上調表達,smpmp3-1受低溫和干旱處理表達上調,而smpmp3-2則下調表達(wangdh等.2013.molecularcloningandexpressionoftwoplasmamembraneprotein3(smpmp3)genesfromsalviamiltiorrhiza(來自丹參的兩種細胞質膜蛋白3(smpmp3)基因的分子克隆和表達).russianjournalofplantphysiology60:155-164.)。玉米有十個rci2基因,其表達量均受干旱脅迫影響,其中zmrci2-8表達量顯著下調(zhaoy等.2014.identificationandcharacterizationoftherci2genefamilyinmaize(zeamays)(玉米(zeamays)中rci2基因家族的鑒定和特性).journalofgenetics93:655-666)。玉米是我國最主要的糧飼兩用作物之一,磷對玉米早期生長發育起關鍵性作用,因此種植玉米的地方經常需要補充大量的磷肥,耗費了大量的人力和物力。另外,土壤鹽漬化是一個世界性的資源和生態問題,我國有鹽堿地區3.6×107hm2,約占總耕地面積的十分之一,由于灌溉不當和大量施用化肥等原因,次生鹽堿化土壤面積還在繼續擴大,從而導致可耕地面積逐年下降,有研究指出在未來的25年內耕地面積將減少30%,所以土壤鹽漬化嚴重制約著農林業生產和生態環境建設。由于鹽漬化的土壤中含有鈉、鉀、鈣、鎂等的硫酸鹽、氯化物、碳酸鹽和重碳酸鹽,而且含量較高,因此對玉米種子的影響極為嚴重,鹽漬化的土壤會影響玉米種子的早期發芽率及發芽指數,阻礙玉米種子萌發,降低玉米種子中抗氧化酶活力和抗氧化物質含量,影響胞內氧化還原勢以及k+/na+離子平衡,對玉米細胞產生傷害,進而導致玉米種子發芽率極低,甚至不發芽。因此,非常迫切需要提供一種既耐低磷脅迫又耐高鹽脅迫的玉米育種方法。技術實現要素:本發明所要解決的技術問題是植物在低磷條件下生長不良以及高鹽環境下發芽率低的問題以及過度施用磷肥或土壤劣化尤其是導致的問題以及土壤鹽漬化衍生的問題。為了解決上述問題,本發明在第一方面提供了zmrci2-8基因,所述基因具有如seqidno.1所示的核苷酸序列。本發明在第二方面提供了zmrci2-8蛋白,所述蛋白具有如seqidno.2所示的氨基酸序列。本發明在第三方面提供了本發明第一方面上述的基因在提高植物耐非生物脅迫能力中的應用。本發明所述基因可響應多種非生物脅迫,名稱為zmrci2-8,來源于玉米屬的玉米(zeamaysl.)。zmrci2-8有2個外顯子,1個內含子,包含231bp的開放式閱讀框,其序列如參見seqidno.1所示。所述基因編碼的蛋白具有76個氨基酸,含有upf0057保守結構域,有兩個疏水性跨膜結構域,屬于rci2家族,其氨基酸序列如seqidno.2所示。通過對低磷條件下zmrci2-8轉基因植株的研究發現側根數目增加,表明zmrci2-8基因對根系構型有影響。在鹽脅迫條件下轉基因株系的發芽率比野生型的明顯提高,表明zmrci2-8基因參與鹽脅迫調控機制,能夠提高轉基因株系對鹽脅迫的耐受性。因此,本發明為農作物的磷高效吸收利用提供了更有效果的基因資源,在基因工程改良植物的營養高效性能和相應非生物脅迫研究中將發揮重要作用,有望改善植物磷效率,培育磷高效農作物。另外,本發明的所述基因還參與調控養分吸收與葉片光合或蒸騰功能,用于提高植物碳水化合物的積累,提升籽粒營養品質及產量。附圖說明圖1是zmrci2-8基因結構示意圖,玉米zmrci2-8位于第9染色體(chr.9:105271795-105597302bp),包括2個外顯子(以e1和e2黑色方框表示),1個內含子(以兩個外顯子之間的折線表示)。圖2是zmrci2-8跨膜結構示意圖,zmrci2-8的全長cdna為1067bp,orf(開放閱讀框)位于其143-373位堿基,全長為231bp,編碼76個氨基酸。利用tmhmm2.0對其跨膜結構進行預測,結果顯示,該蛋白含有2個跨膜結構域。實線1代表跨膜結構機率;實線2代表細胞膜內機率;實線3代表細胞膜外機率。圖3是玉米zmrci2-8啟動子順式作用元件分析,利用place(http://www.dna.affrc.go.jp/place/signalscan.html)數據庫分析zmrci2-8啟動子順式作用元件,發現zmrci2-8啟動子包含一系列植物逆境脅迫響應元件,包括脅迫誘導響應元件abrelaterd1、磷饑餓響應元件p1bs、干旱、低溫及aba響應元件myc、赤霉素和細胞分裂素響應元件。其中磷饑餓響應元件p1bs存在于多數磷饑餓響應基因中,并且受缺磷誘導的osrci2-8啟動子序列中也存在兩個p1bs順式元件,由此推測zmrci2-8是一個受缺磷誘導響應基因。圖4是玉米zmrci2-8蛋白與擬南芥atrci2(a-h)、水稻rci2之間的進化關系。圖5是正常營養條件,玉米自交系b73抽雄期zmrci2-8的組織表達特征。田間養分充足條件下,玉米自交系b73抽雄期收獲營養器官(根、節、節間、葉片)和生殖器官(雌穗、雄穗),以根中zmrci2-8表達水平為參照,發現各組織中zmrci2-8表達水平由高到低依次為葉、節、根、節間、雄穗、雌穗。葉片中zmrci2-8表達水平是根的4.5倍,節中水平比根高1.9倍。雌穗和雄穗中zmrci2-8表達水平僅及根中水平10%。玉米抽雄期zmrci2-8在葉、根中表達量較高。其中,根和葉對養分和水分的吸收或代謝具有重要作用,zmrci2-8表達量高低影響了葉片蒸騰強度和根系吸收功能。圖6是gfp-zmrci2-8融合蛋白的亞細胞定位,分別為egfp空載熒光信號和zmrci2-8-egfp熒光信號。從圖中可以看出,zmrci2-8在質膜中表達,與signalp4.1server和wolfpsort軟件預測結果相符合。圖7是zmrci2-8根中的表達特征(在缺氮、磷、或鉀10d(天)后),可以看出,低氮、低磷、低鉀處理玉米b73十天后,zmrci2-8表達只受低磷誘導響應強烈,在根和地上部均顯著性上調。低磷條件下根中zmrci2-8相對表達量較正常營養條件上調32.6倍,低氮或低鉀下,zmrci2-8表達量無顯著差異。圖8是zmrci2-8在地上部氮、磷、鉀養分脅迫10d條件下的表達特征(在缺氮、磷、或鉀10d后),可以看出,玉米b73自交系地上部zmrci2-8的相對表達量比正常營養上調31.2倍,低氮或低鉀條件下zmrci2-8表達量較正常營養無顯著差異。結果表明,zmrci2-8基因在玉米根和地上部中均表達,并且其表達受低磷特異性誘導上調。圖9是zmrci2-8超表達株系表型分析(土培12dcol-0和超表達株系l10、l12、l16的整體長勢,圖的比例尺為1cm),篩選轉基因株系至t3代得到純合株系,t3代轉基因純合株系進行土培并觀察其表型。將col-0作為對照,觀察不同生長時期擬南芥轉基因株系的表型。通過前期土培觀察,發現l10、l12、l16與col-0相比蓮座葉數量、葉片大小、葉色、葉形、抽薹時間均無明顯差別。圖10是zmrci2-8超表達株系表型分析(土培27dcol-0和超表達株系l10、l12、l16的整體長勢,圖的比例尺為1cm)可以看出,移苗后土培27d,轉基因株系l10、l12、l16與col-0相比株高無明顯差別,開花時間、花序大小基本一致。圖11是zmrci2-8超表達株系表型分析(土培47dcol-0和超表達株系l10、l12、l16的整體長勢,圖的比例尺為1cm),轉基因株系l10、l12、l16與col-0相比株高高度基本一致。zmrci2-8-10平均株高17.5cm,l12平均株高17.6cm,l16平均株高17.1cm,col-0平均株高17.9cm。與col-0相比,轉基因株系l10、l12、l16的果莢大小、長短無明顯差異。col-0、l10、l12、l16整株植株的平均分枝數分別為2.4、2.3、2.4和2.3個,各株系分枝數相近,無明顯差異。表明在正常營養土培條件下轉基因株系生長發育、開花結莢與col-0均無明顯差異,可以推測正常營養條件zmrci2-8對擬南芥生長發育無顯著性影響,但是zmrci2-8表達受低磷強烈誘導。圖12是zmrci2-8轉基因株系缺磷培養7d后的主根長(l10、l12、l16代表獨立的zmrci2-8轉基因株系),可以看出,轉基因株系的主根長在正常營養條件以及缺磷條件下與對照col-0都不存在差異。在缺磷條件培養7d后,超表達株系的主根長較col-0沒有顯著差異。圖13是zmrci2-8轉基因株系缺磷培養7d后的可見側根數(l10、l12、l16代表獨立的zmrci2-8轉基因株系),對其可見側根數進行統計發現,缺磷處理后這三個轉基因株系的可見側根數與col-0相比都存在顯著差異。側根數量的增多是植物響應低磷的表型。由此可以推斷,可能zmrci2-8主要對側根產生影響,zmrci2-8可能通過調控側根發育影響磷脅迫。圖14是zmrci2-8轉基因株系和col-0不同濃度nacl處理條件下第三天的發芽率。l10,l12,l16代表獨立的zmrci2-8轉基因株系。“基因表示處理間存在顯著差異(p<0.05)。120mmol/lnacl濃度下,轉基因株系l10、l12、l16的發芽率明顯高于col-0,150mmol/lnacl濃度下,轉基因株系的發芽率差異較大,轉基因株系l12發芽率基本與col-0無明顯差異,但株系l16的發芽率仍顯著高于col-0。由此可以推測出zmrci2-8對擬南芥耐鹽脅迫有影響。圖15為載體pezs-nl結構示意圖,其中“camv35promoter”表示camv35啟動子,“ampicilinresistance”表示氨芐青霉素抗性基因。具體實施方式如上所述,本發明在第一方面提供了zmrci2-8基因,所述基因具有如seqidno.1所示的核苷酸序列。本發明在第二方面提供了zmrci2-8蛋白,所述蛋白具有如seqidno.2所示的氨基酸序列,其中在第2至3個氨基酸位置處的氨基酸為均為ser(絲氨酸)。本發明在第三方面提供了本發明第一方面上述的基因在提高植物耐非生物脅迫能力中的應用。本發明人利用place數據庫分析zmrci2-8基因啟動子順序作用元件驚訝地發現,該基因啟動子區域存在myc順式元件和pibs順式元件,myc的核心序列為canntg,并且實驗證實可用于多種抗逆基因啟動子上。p1bs順式元件的核心序列為gnatatnc,并且實驗證實可用于磷誘導基因啟動子上。在一些優選的實施方式中,所述非生物脅迫為低磷脅迫和/或高鹽脅迫。在一些優選的實施方式中,所述低磷脅迫為種植基質中的磷濃度為不大于2.000×10-3mol/l,優選為2.000×10-3mol/l至5.000×10-9mol/l。在一些優選的實施方式中,所述高鹽脅迫為種植基質中的鹽濃度為不低于120mmol/l,優選為120mmol/l至150mmol/l。在一些優選的實施方式中,所述植物在低磷脅迫條件下側根生長得到促進;和/或所述植物在高鹽脅迫條件下種子發芽率得到提高。所述基因提高植物耐受低磷脅迫的能力可能通過提高磷的利用率以及對磷更加有效的調控來實現。在一些實施方式中,其中將權利要求1所述的基因轉入植物中以提高所述基因在植物體內的表達,例如提高所述基因在根或葉片中的表達。本發明人驚訝地發現,本發明的所述基因不僅能夠提高植物的耐低磷脅迫能力和耐高鹽脅迫能力,而且還在養分吸收、光合作用、蒸騰作用、碳水化合物積累、籽粒營養品質、生物量、籽粒產量等方面發揮作用。因此,本發明的所述應用還可以進一步包括本發明第一方面所述的基因在如下方面中的應用:(1)調控養分吸收能力、光合作用或蒸騰功能;(2)提高碳水化合物積累;(3)提升籽粒營養品質;和/或(4)提高生物量和/或籽粒產量。本發明人通過分析定量pcr結果發現,zmrci2-8在玉米的地上部和根部都受缺磷強烈誘導,并且根部上調倍數較高。對抽雄期后一周玉米各組織zmrci2-8表達量分析可知,zmrci2-8的表達量由高到低依次是葉、節、根、節間、雄穗和雌穗,葉片和根中的表達量明顯多于其它組織中的表達量。由此推測zmrci2-8可能在玉米葉片光合作用或水分、養分轉運過程中起到一定的作用。在一些優選的實施方式中,所述植物為禾本科植物或豆科植物,優選為玉米(zeamaysl.)或擬南芥。本發明的所述zmrci2-8基因進行亞細胞定位,發現其在質膜表達,符合rci2家族基因的定位預測。以下實施例進一步說明本發明的內容,但不用來限制本發明所要保護的范圍。下述實施例中所用方法如無特別說明,均為常規方法。材料的準備1、植物材料、菌株與載體本發明中所用到的材料包括:玉米(zeamaysl.)材料為自交系b73;擬南芥材料為野生型哥倫比亞col-0;大腸桿菌dh5α菌感受態細胞(codeno.cb101,天根生化科技有限公司);農桿菌感受態細胞gv3301(購自天根生化科技有限公司);zmrci2-8基因ta克隆載體19-t載體(codeno.d102,takara公司);zmrci2-8擬南芥超表達載體psuper1300+-kanamycin(上海北諾生物科技有限公司);zmrci2-8轉洋蔥表皮細胞亞細胞定位載體pezs-nl(參見圖15)。2、生化試劑與試劑盒ms培養基(phytotech公司);primestarhsdnapolymerase(takara公司);taq酶,2×pcrmix(北京匯天東方生物技術公司);rana(北京匯天東方生物技術公司);dnamarkers、rnasea(北京天根生物技術公司);t4dna連接酶、限制性內切酶、sal-hf、bamh1-hf(neb公司);oligod(t)18、反轉錄酶、dna酶、rnase抑制劑(invitrigen公司);sybegreen(定量pcr試劑,takara公司);卡那霉素(100mg/ml),利福平(25mg/ml),氨芐青霉素(100mg/ml),潮霉素(50mg/ml)等抗生素(bbi公司);其他化學試劑均為分析純試劑(北京化學試劑廠);普通瓊脂糖凝膠dna回收試劑盒、質粒小提試劑盒(北京天根生物技術公司);trizol試劑盒(invitrigen公司);其他化學試劑均為分析純試劑(北京化學試劑廠)。3、pcr擴增引物4、培養基配方(1)玉米營養液配方注:將各營養液成分按表所示,用1mol/lnaoh調節ph至6.0。(2)1/2ms培養基(固體100ml):ms0.217g,蔗糖3g,瓊脂粉0.8g,用1mol/lkoh調節ph至5.8,121℃高溫滅菌20min(分鐘)。(3)擬南芥缺磷培養基(固體)注:將各營養液成分按表所示,加入1%的蔗糖,用1mol/lkoh調節ph至5.8±0.1成后加入1%瓊脂。(4)擬南芥鹽脅迫培養基(固體):將不同濃度的nacl溶液(0mmol/l、120mmol/l、150mmol/l)添加到1/2ms培養基。(5)lb培養基(固體100ml):胰蛋白胨1g,酵母提取物0.5g,nacl1g,瓊脂粉1.5g,121℃高溫滅菌20min。(6)液體培養基:除不加瓊脂粉外,其余與1/2ms或lb固體培養基配方相同。(7)篩選培養基:1/2ms或lb固體培養基高壓滅菌后待溫度降至50℃左右時加入抗生素,搖勻后鋪平板。實施例1、供試材料培養方法1、擬南芥培養及轉基因幼苗篩選種子春化:取適量擬南芥種子于1.5ml離心管中,用去離子水沒過所有種子,置于4℃冰箱春化2天。種子消毒:向裝有種子的1.5ml離心管中加入75%乙醇,滅菌1min,滅菌水沖洗一遍,再加入2%次氯酸鈉滅菌2min,用滅菌水沖洗5-7遍。點種及培養:用10培養移液槍將種子均勻點在1/2ms+50mg/ml潮霉素的培養基上,將點有種子的培養基置于培養室(光周期:16h白天/8h黑夜、溫度22/20℃、濕度100%)中培養。將幼苗移栽至花盆中,盆中裝有滅過菌的營養土(蛭石/營養土的體積比=1:1),移栽后在花盆表面覆蓋一層保鮮膜,每5天澆400ml水,10天后揭開。揭去保鮮膜后,每3天澆400ml水。轉基因幼苗篩選:取t0代擬南芥種子春化、消毒,將種子均勻點在1/2ms+50mg/ml潮霉素的培養基上。培養皿外包裹一層黑色塑料膜,置于培養室黑暗培養5-7天,幼苗莖抽出很高的幼苗視作具有抗性的轉基因幼苗,從具有抗性的轉基因植物中每個株系挑選13株幼苗,編號,移至1/2ms培養基培養3-5天,待其長出4片幼葉后移至營養土中培養,完成其整個生育期。收獲種子繼續篩選至t3代獲取純合轉基因株系。2、玉米培養及供試樣品獲取方法玉米水培培養:挑選籽粒飽滿、大小基本一致的玉米種子,用去離子水沖洗3-5遍,再用10%h2o2浸泡30min,去離子水水沖洗干凈,飽和caso4溶液中浸泡5-6h,移至鋪有濾紙的托盤中,用去離子水將濾紙潤濕,胚乳面向下擺放到濾紙上,上層鋪濕潤濾紙,用黑塑料遮蓋托盤,避光催芽。每天早中晚各噴水一次,保持濾紙濕潤。待芽長到1-2cm時,用濕潤濾紙卷苗,直立放到盛有去離子水的水培罐中,黑塑料遮蓋培養。當子葉萌出后,揭去黑塑料,給予適當光照。玉米長至兩葉一心時,精選出苗整齊的幼苗去掉胚乳,移栽至各處理營養液中生長。取樣:低氮、低磷、低鉀表達特征樣品,玉米自交系b73對照、低氮、低磷、低鉀營養液處理,取樣1次,取樣部位為移栽后10d幼苗的地上部分和根。組織表達特征樣品玉米自交系b73大田條件下抽雄期,取樣1次,取樣部位為根、節、節間、雌穗、葉和雄穗。樣品取自大田條件下玉米自交系b73品種。將收集的樣品用液氮迅速冷凍之后存放于-80℃冰箱。實施例2、目標基因片段克隆1、目標基因反應系統反應程序:98℃、5min,(98℃、30s,60℃、30s,72℃、20s)30循環(cycle),72℃、10min。反應結束后,取反應液進行1%瓊脂凝膠電泳,之后將擴增所得產物進行膠回收,膠回收步驟參照天根離心柱型試劑盒膠回收。2、重組質粒載體構建加polya尾巴,200μlep管,10μl體系:dna7μl、taq緩沖液1μl、taqdnase1μl、dntps1μl。混勻后,72℃下孵育30min。t載體19-tvector連接,200μlep管,5.0μl體系:dna2μl、t載體0.5μl、溶液ⅰ2.5μl。常溫連接約3h。連接目的載體,連接t載測序正確后進行膠回收,構建重組質粒,常溫連接3h。反應體系:10×連接酶緩沖液0.5μl、目的基因a、載體(表達載體)b、t4dna連接酶0.5μl、滅菌ddh2o補足至5μl。3、目標基因檢測及回收將連接產物轉化進入大腸桿菌感受態細胞dh5α后,涂布在lb+抗生素培養基平板上,倒置放于37℃恒溫培養箱中12-16h。在超凈臺中挑單菌落作為cdna,進行pcr擴增。將陽性克隆菌液加入lb+抗生素液體培養基中活化12h。吸取1-1.5ml菌液,參照天根質粒小提試劑盒(購自天根生化科技有限公司)說明,提取質粒。酶切檢驗/膠回收,酶切檢驗使用10μl反應體系,檢驗成功菌液送去測序;酶切片段回收則使用50μl反應體系,膠回收目標片段。反應體系如下:10×neb緩沖液1μl5μldna≤0.5μg≤2μg100×bsa0.1μl0.5μl內切酶10.2μl1μl內切酶20.2μl1μl總共10μl50μl實施例3、植物dna、cdna的提取及檢驗1、ctab法提取擬南芥t0代葉片dna取0.5g新鮮植物葉片置于研缽中,加入適量液氮,迅速研磨成均勻粉末。將粉末移入1.5ml離心管,加入800μlctab分離緩沖液,上下顛倒離心管,混合均勻。將盛有樣品的離心管置于65℃水浴中保溫30min,其間每隔3-4min輕搖混勻。低溫12000rpm(轉/分鐘)離心5min,將上層水相轉移至新的離心管中。加入等體積的tris-酚和氯仿異戊醇,上下顛倒離心管,混合均勻;低溫12000rpm離心5min,將上層水相轉移至新的離心管中。加入等體積的氯仿異戊醇上下顛倒離心管,混合均勻;低溫12000rpm離心5min,將上層水相轉移至新的離心管中。加入0.6倍體積的異丙醇,輕輕混勻,置于-20℃沉淀30min。低溫12000rpm離心5min去上清,加入500μl70%乙醇洗滌沉淀,常溫12000rpm,2min,去上清,沉淀盡量干燥。加入適量含有rnase的te(20μl)溶解沉淀,37℃水浴30min,消化rna。取適量基因組dna樣品,電泳檢測。2、trizol法提取玉米總rna研磨制樣:先向研缽中加入少量液氮,等液氮揮發使研缽預冷。再向研缽內倒入適量液氮。從-70℃冰箱取出組織,用鑷子從凍存管中夾取一小塊放入研缽中。迅速研磨組織塊,成粉末時加入1ml的trizol,繼續研磨,可看到粉末成漿,由濃變稀,直至組織樣全部溶解,用移液槍吸到無酶ep管。trizol法提取rna:室溫靜置3-10min,使蛋白質變性;加入1/4-1/5體積的氯仿,顛倒震蕩1min,室溫靜置3-5min,靜置分層,然后4℃離心,12000rpm,5min。吸取上清至無酶的ep管中,加入等體積的酚:氯仿,顛倒震蕩1min,室溫靜置3-5min,靜置分層,然后4℃離心,12000rpm,5min,取上清。若是細胞rna提取可只加等體積氯仿抽提一次,但對于組織一般都要重復該步驟,至沒有蛋白層為止,一般抽提兩次。加入等體積的預冷的異丙醇,顛倒混勻,冰上靜置30min,然后4℃離心12000rpm,30min,棄上清。加入100ul70%乙醇,將沉淀吹起來,注意不要吹散,然后4℃12000rpm,5min。棄上清,空氣中揮發乙醇,至壁上無液滴。加入適量depc水,(一般為20μl)溶解。分光光度計檢測rna濃度和od值,將totalrna稀釋至1μg/μl,備用。3、植物總rna純化取5-10g樣品放入研缽中,加入過量液氮,迅速研磨成均勻的粉末(在研磨過程中,保證樣品始終浸在液氮中,研磨約30min)。待液氮自然揮發干凈后,將樣品轉入100ml的離心管中,加入6-8倍體積的裂解液1,輕輕攪拌后,0℃冰浴20min。12000rpm,4℃離心15min。取上清,加入1/3體積的3mol/l醋酸鈉,1/5的氯仿異戊醇,輕輕搖勻,0℃冰浴20min。2000rpm,4℃離心15min(加5倍體積裂解液2溶解沉淀以提取dna)。取上清,加入等體積冰浴冷卻的異丙醇,混勻且冰浴10min。5000rpm,4℃離心10min,將沉淀溶于5ml含有0.1%sds的te緩沖液中,加入8mol/llicl使終濃度至2.5mol/l,冰浴3h或4℃過夜。12000rpm,4℃離心10min。70%乙醇洗滌沉淀兩次,空氣干燥10min,溶于無菌水(depc焦碳酸二乙酯,處理),加入1/10體積10%的sds,重復第4~6步驟,進一步除去蛋白質,70%乙醇20℃長期保存。4、rna反轉錄:反應體系如下:dntp混合液(各為10mmol/反應)1μl、oligodt引物(寡聚胸腺嘧啶引物)(2.5μm)1μl、總rna5μl,用ddh2o補足至10μl。反應程序:65℃、5min。實施例4、轉基因植株構建1、制備農桿菌感受態細胞挑取少許農桿菌lba4404,接種于5mllb液體培養基(含50mg/lstr)中,28℃、200rpm培養過夜。取2ml培養物于lb液體培養基(含50mg/lstr)中繼續培養,直至od800為0.5左右。將培養物置冰浴中30min,4℃、5000rpm、離心5min,棄去上清液。用10ml冷的0.1mol/lnacl懸浮菌液。4℃、5000rpm,離心5min,棄去上清液。用1ml冷的cacl2(20mmol/l)懸浮,分裝成50μl/管,液氮中冷凍后至-80℃保存。2、農桿菌電轉化將20μl農桿菌和0.5μl質粒混勻;用槍加入4mm電轉化杯中,蓋上蓋,貼標簽;在電擊器上2500v/6ms進行電擊,促使質粒進入農桿菌;電擊后,用槍將液體移到裝有500μllb培養液的2ml離心管中;寫標簽,用石蠟膜封口;在28℃生化培養箱中搖晃培養4h左右,讓菌在有氧條件下復蘇;將離心管進行離心1min,使菌體沉降到管壁上;準備lb平板,寫標簽,用酒精擦拭三角玻璃棒,然后在火焰上燒10s左右滅菌,放置冷卻備用;用槍吸取100μl上清液,將剩下液體倒掉,在將槍中液體打入離心管中,并用槍反復吸打,沖下壁上菌體混勻;吸出菌液,滴在平板上,用玻棒涂抹均勻;蓋上蓋,用石蠟膜封口,放入28℃生化培養箱中48h(2天),即可長出菌落。3、擬南芥蘸花轉化將低溫保存的帶有表達載體的農桿菌菌株,分別在對應抗性平板上劃線,挑取單菌落接種于5ml的已加入對應抗生素的液體lb培養基中,28℃下250rpm振蕩培養18-24h;然后相同條件下按照1:100接種擴大培養,總菌液體積為200ml,直到od600值在0.8-1.0范圍內;離心沉淀并去上清液,用等體積的5%的蔗糖溶液重懸菌體,在菌液中加入0.02%-0.04%的silwetl-77混勻;將擬南芥的花序在菌液中蘸取0.5-1min即可(在轉化前要先將擬南芥的角果剪掉);然后將擬南芥苗放置于黑暗,濕潤的條件下16h,一周后,重復侵染一次;收種后種植后代,向12d的幼苗上噴灑54mg/ml除草劑或在將種子播種于抗性ms平板上以篩選陽性植株。實施例5、基因槍操作及共聚焦顯微鏡觀察1、材料:洋蔥內層表皮2、試劑:2.5mol/lcacl2,0.1mol/l亞精胺,無水乙醇3、前期準備:撕取洋蔥內層表皮大約2×2cm,將撕取的洋蔥表皮組織置于現配的1/2ms培養基上,黑暗培養12h。4、制備dna-金粉復合體:配置2.5mol/lcacl2和0.1mol/l亞精胺各1ml。將金粉渦旋震蕩5min,使其在無菌水中呈懸浮狀態。吸取金粉懸浮液8.5μl于1.5mlep管中,并順序加入5μg質粒dna,17μlcacl2和8μl亞精胺。震蕩5min,冰上靜置10min,3000rpm離心1min,棄上清。加入100μl無水乙醇,不破壞沉淀洗滌,棄上清,重復2次。加入10μl無水乙醇,備用。5、基因槍步驟:取10μl制備好的dna-金粉復合體,均勻涂布在轟擊膜中間孔徑處,基因槍上方放置壓力膜在孔內轟擊材料,在艙內壓力降為0時開艙門取出材料。6、培養:轟擊完畢將洋蔥表皮置于1/2ms培養基中,28-30℃培養箱黑暗培養22-24h,進行共聚焦顯微鏡觀察。結果本發明以玉米自交系b73為材料,運用分子生物學技術,對zmrci2-8的序列特征、進化特點、表達特征和在細胞中的表達部位、植物生長發育過程中的作用進行了初步研究,主要結果如下:(1)zmrci2-8含有upf0057保守結構域,兩個跨膜結構域,屬于rci2家族(參見圖2和4)。(2)通過分析定量pcr結果發現,zmrci2-8在玉米的地上部和根部都受缺磷強烈誘導,并且根部上調倍數較高。對抽雄期后一周玉米各組織zmrci2-8表達量分析可知,zmrci2-8的表達量由高到低依次是葉、節、根、節間、雄穗和雌穗,葉片和根中的表達量明顯多于其它組織中的表達量。推測zmrci2-8基因可能影響葉片的光合作用和根系建成及養分吸收(參見圖5)。(3)對zmrci2-8進行亞細胞定位,發現其在質膜表達,符合rci2家族基因的定位預測(參見圖6)。(4)低磷條件下,轉基因株系可見側根數顯著增多,表明zmrci2-8基因響應磷饑餓誘導(參見圖7、8、11、13)。(5)鹽脅迫條件下轉基因株系的發芽率比野生型的明顯提高,表明zmrci2-8基因參與鹽脅迫調控機制,能夠提高轉基因株系對鹽脅迫的耐受性(參見圖14)。(6)正常條件下培養,轉基因植株的整體長勢例如蓮座葉數量、葉片大小、葉色、葉形、抽薹時間、主根與對照植株沒有明顯差別(參見圖9、10和12)。討論通過序列分析和對其跨膜結構預測,發現zmrci2-8基因有2個外顯子,1個內含子,包含231bp開放式閱讀框,編碼76個氨基酸,含有兩個疏水性跨膜結構域,屬于rci2家族。受低磷脅迫后zmrci2-8的表達量顯著上調,根部的上調倍數略高于地上部,說明zmrci2-8在轉錄水平上受缺磷的強烈誘導。玉米b73抽雄期的各組織中,zmrci2-8在營養器官中的表達量較高,其中葉片的表達量高于其它各組織,說明zmrci2-8還在玉米葉片光合作用或水分、養分轉運過程中起到一定的作用。利用農桿菌蘸花轉化法得到擬南芥轉基因株系,正常營養土培條件下,轉基因株系的表型與野生型基本一致,在抽薹時間、開花時期、衰老程度等不同生長發育時期均無明顯差異。這一結果表明轉基因植株的正常生長并沒有受到不利影響。分析zmrci2-8基因啟動子順式元件發現,啟動子區域存在myc順式元件和pibs順式元件,myc的核心序列為canntg,可用于多種抗逆基因啟動子。p1bs順式元件的核心序列為gnatatnc,可用于磷誘導基因啟動子。為探究zmrci2-8基因啟動子中myc順式元件和pibs順式元件是否對鹽害及磷饑餓誘導表達,做了低磷和鹽害處理試驗。從低磷處理可以看出,zmrci2-8轉基因植株在低磷條件側根數目增加,說明zmrci2-8基因對根系構型有影響。鹽脅迫處理試驗結果可以發現,zmrci2-8還提高了植株對鹽脅迫的耐受性。序列表<110>中國農業大學<120>zmrci2-8基因及其在促進非生物脅迫條件下植物發芽和側根生長中的應用<160>12<170>patentinversion3.5<210>1<211>1067<212>dna<213>玉米(zeamaysl.)<400>1gtaaacgcacccatcggcatcttcccatggcagccagcgtcggattccgcatcattccct60cgcccagttgtcccccccgtacaaattaacacctaagctaaccttccgggctagcttgtg120tgtgtggcctcggaggtccgggatgagctccggcggctgctcgacgtgcctcgaggtcat180cttcgccgccatcctcccgccgctcggcgtcttcttccgctacggctgctgcagctccga240gttcttcgtctccctcctcctgacgctgctgtgctacgtccccggcgtcgcgtactccct300ctacgtcatcctccggacgccgcccgagccgccgggcatcgacggggagcggccctacga360catgctcgcctgacggctttgcttgtacttgtacagctccacccaacaacaacaacgcgt420aggcctgtgcaatgcaacacacactcgctgtatgtagtagctgtaggtagcaggtagcgg480ctcaggacattcctgactgtgttctccagcgtcacgtacgtccctgttcctcgtcgcgtg540tgatgactgtgcaattgttggtcattggtttgctgattctttcgatcagtttatcacgaa600cagtagggtgctgccgtgctggactgctgctcgtctgtttgccgatcacgcagcagttgg660tggtaattggcaactgatataaaatttatgcgtgtactcatgtataaatcccaaccagta720atgcggttggaattaaaagaaaggtattgtttggatatggtttacggctacgctattccc780actaaggtcttgttcggttaatcccattacccatggattgtacgggattggaaaaattta840agaagaagtttgacttgcttgggattcaaacccatccaatcccactcaatccacatggat900tgggagctaaccgaacaagccctaataatgatatttgtagacaccgttaggtcttcccat960tcgcattcgcatgtggatcggataggacctggggtgtgggggtggggcataattagggac1020acccgaattactccttaacaaacgcatttagagaaataagaattaaa1067<210>2<211>76<212>prt<213>玉米(zeamaysl.)<400>2metserserglyglycysserthrcysleugluvalilephealaala151015ileleuproproleuglyvalphepheargtyrglycyscysserser202530gluphephevalserleuleuleuthrleuleucystyrvalprogly354045valalatyrserleutyrvalileleuargthrproproglupropro505560glyileaspglygluargprotyraspmetleuala657075<210>3<211>26<212>dna<213>人工序列<400>3gggcccatgagctccggcggctgctc26<210>4<211>28<212>dna<213>人工序列<400>4actagttcaggcgagcatgtcgtagggc28<210>5<211>21<212>dna<213>人工序列<400>5cgaggtcatcttcgccgccat21<210>6<211>18<212>dna<213>人工序列<400>6ccgctccccgtcgatgcc18<210>7<211>26<212>dna<213>人工序列<400>7gtcgacatgagctccggcggctgctc26<210>8<211>26<212>dna<213>人工序列<400>8ggatccggggcgagcatgtcgtaggg26<210>9<211>24<212>dna<213>人工序列<400>9taagctgccgatgtgcctgcgtcg24<210>10<211>27<212>dna<213>人工序列<400>10ctgaaagacagaacataatgagcacag27<210>11<211>20<212>dna<213>人工序列<400>11agaggttgacgagcagatga20<210>12<211>20<212>dna<213>人工序列<400>12cctcttcttcctcctcgtac20當前第1頁12
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1