一種提高spCEMA對植物病原菌抗性的方法及轉基因煙草材料與流程
本發明涉及一種提高spcema對植物病原菌抗性的方法,還涉及spcema::erbd的植物表達載體。屬于植物基因工程技術領域。
背景技術:
在農業生產中,植物病害尤其是真菌病害一直是限制農作物產量提高的重要因素。長期以來,廣泛采用的防治手段主要為化學防治,但由于病原菌變異速度快,會對各種化學農藥產生耐藥性或抗藥性,同時農藥所帶來的環境污染和生態平衡問題也不容忽視。因此,為保障食品安全,減少化學防治方法對環境的破壞,急需開發高效并對環境安全的植物病害防治手段。
近年來,基因工程在植物抗病的研究上已經取得了初步的進展。研究表明,在植物中表達不同來源的抗菌肽基因能顯著增強植物對病原菌的抗病性(gaoetal.,2000;jaynesetal,1993;jhaetal.,2009)。但利用抗菌肽獲得的轉基因植物材料抗病能力較差,還不能滿足生產的的需要。為獲得高效的植物抗病基因,除繼續在不同物種中篩選、克隆抗菌肽基因外,利用分子進化手段對抗菌肽進行定向改良,在現有基礎上提高其抗菌效果是一條較為有效的途徑。現有研究發現,不同的抗菌肽具有不同的抑菌機制,包括結合并破壞真菌細胞膜,穿過細胞膜進入細胞中抑制不同的靶標等。因此,與真菌細胞膜的互著是抗菌肽發揮抑菌活性的重要步驟。我們設想,如能增加抗菌肽在真菌細胞膜上的富集能力,將提高抗菌肽的抑菌效果。在真菌中,麥角甾醇是細胞膜上獨特的脂類成分,能夠調控真菌細胞膜的流動性和滲透性,是細胞生存的關鍵因子(sanglardetal.,2003)。cema是由殺菌肽ceropina的n-端8個氨基酸和經改造的蜂毒素c-末端組成的陽離子抗菌肽,我們實驗室進一步在其n端添加信號肽獲得了spcema(牛國清等,2002)。將spcema導入植物中,可提高植物抗病性,但提高幅度仍有限。在本發明中,以真菌細胞膜上的麥角甾醇為靶標,利用分子改良手段在現有抗菌肽spcema上添加麥角甾醇結合域erbd(luccaetal.,1998),提高對植物病原真菌的靶向性和富集能力,從而達到理想的抗菌效果。
技術實現要素:
針對現有技術存在的上述不足,本發明的目的在于提供一種提高spcema對植物病原菌抗性的方法,解決目前植物抗病基因工程中抗菌肽的抑菌效果不理想的問題,利用分子改良手段在spcema的c末端融合麥角甾醇結合域erbd,提高抗菌肽在植物病原菌細胞膜上的靶向性和富集能力,從而進一步增強轉基因植物對病原菌的抗性。
本發明還提供該融合抗菌肽基因的植物表達載體及轉基因煙草材料。
為實現上述目的,本發明采用如下技術方案:一種提高spcema對植物病原菌抗性的方法,以植物病原真菌細胞膜上的麥角甾醇為靶點,利用分子改良手段在spcema的c末端融合麥角甾醇結合域erbd,并獲得相應的融合抗菌肽基因;如seqidno.1所示的核苷酸序列和seqidno.2所示的氨基酸序列,構建植物表達載體。
進一步,具體步驟為:由于麥角甾醇結合域erbd和spcema的分子量小,采用人工合成的方式(常州基宇生物科技有限公司)獲得兩端分別添加bamhi與ecori酶切位點、中間用linker序列連接的融合抗菌肽,并連接于大腸桿菌克隆載體puc57上,命名為puc57-spcema::erbd。隨后,bamhi與ecori酶切puc57-spcema::erbd,回收spcema::erbd片段,并與植物表達載體進行連接,獲得正確的spcema::erbd植物表達載體。
一種融合抗菌肽基因的轉基因煙草材料,利用構建的上述植物表達載體,經遺傳轉化后導入野生型煙草中組成性表達。
相比現有技術,本發明具有如下有益效果:
1、本發明利用分子改良手段在spcema的c末端融合了麥角甾醇結合域erbd,并獲得一個新的融合抗菌肽基因。與野生型抗菌肽相比,改良后的抗菌肽能更加有效的抑制植物病原真菌的萌發及生長。
2、本發明將融合抗菌肽基因導入野生型煙草中,獲得相應的轉基因煙草。結果顯示,相比野生型spcema,獲得的spcema::erbd轉基因煙草對植物病原菌的抗性顯著提高。表明經分子改良后的spcema::erbd在植物抗病基因工程中具有較好的應用前景。
3、隨著轉基因技術的發展,越來越多的抗菌肽已被應用到植物抗病基因工程中。然而,天然抗菌肽僅有限提高轉基因植株對病原菌的抗性,還不能滿足生產需要。因此,植物抗病基因工程的關鍵是獲得高效、特異的抗病基因。本發明中利用植物病原菌細胞與植物細胞在結構及組成成份上存在差異,以植物病原真菌細胞膜上特有的麥角甾醇為靶點,通過定向改良手段在spcema的c末端融合麥角甾醇結合域erbd,獲得相應的融合抗菌肽,提高其對植物病原真菌的靶向性和富集能力,在相同濃度下,增強抗菌肽對病原真菌的抑制效果,同時減少對非靶標細胞的危害。
附圖說明
圖1spcema::erbd植物表達載體的酶切驗證;
圖中m2000表示marker2000(bioer公司)m15表示marker15(fermentas公司);泳道1-5表示不同spcema::erbd植物表達載體轉化子經限制性內切酶bamhi和ecori酶切的結果。
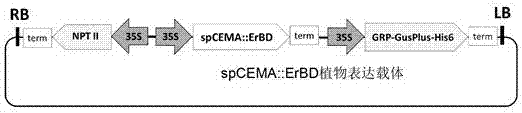
圖2spcema::erbd植物表達載體圖譜;
其中grp-gusplus-his6代表gusplus報告基因,該基因在n端融合了grp信號肽,在c端融合了his6序列標簽;nptii代表新霉素磷酸轉移酶基因,具有卡那霉素抗性;camv35s:來源于花椰菜花葉病毒的植物組成性啟動子;lb:t-dna左邊界;rb:t-dna右邊界。
圖3轉基因煙草的gus染色;
transgenic代表的是轉基因煙草的gus染色結果;non-transgenic代表的是非轉基因煙草的gus染色結果。
圖4轉基因煙草表達量篩選;
橫坐標表示不同轉基因煙草的轉化子,其中spcema::erbd-4和spcema::erbd-42代表的是spcema::erbd轉基因株系,spcema-44和spcema-46代表的是spcema轉基因株系,wt表示野生型煙草植株;縱坐標表示不同植株中外源基因的相對表達水平。
圖5轉基因煙草葉片離體抗病性比較;
a:轉基因煙草葉片對大麗輪枝菌的抗性比較;b:轉基因煙草葉片對煙草赤星病的抗性比較。wt表示野生型煙草葉片;spcema表示spcema轉基因煙草葉片;spcema::erbd表示spcema::erbd轉基因煙草葉片;water表示經水處理的野生型葉片。
圖6轉基因煙草葉片總蛋白提取液對油菜黑斑病原菌的抗性比較;
wt表示野生型煙草葉片總蛋白提取液;spcema表示spcema轉基因煙草葉片總蛋白提取液;spcema::erbd表示spcema::erbd轉基因煙草葉片總蛋白提取液;kpb表示磷酸鹽緩沖液。
具體實施方式
下面結合具體實施例和附圖對本發明作進一步詳細說明。
本發明提供一種提高spcema對植物病原菌抗性的方法,利用分子改良手段將麥角甾醇結合域erbd,融合到spcema的c末端,其具有如seqidno.1所示的核苷酸序列和如seqidno.2所示的氨基酸序列。
【實施例1】spcema::erbd基因的獲取
由于麥角甾醇結合域erbd和spcema的分子量小,采用人工合成的方式(常州基宇生物科技有限公司)獲得兩端分別添加bamhi與ecori酶切位點、中間用linker序列連接的融合抗菌肽,如序列1所示1-165bp為spcema序列;166-189bp為linker序列;190-240bp為erbd序列;241-243bp為終止密碼子序列,如序列2所示1-55aa為spcema編碼;56-63aa為linker編碼;64-80為erbd編碼。并連接于大腸桿菌克隆載體puc57上,命名為puc57-spcema::erbd。
【實施例2】spcema::erbd植物表達載體的構建
為驗證通過分子改良后的spcema::erbd能否進一步提高植株的抗病能力,于是構建了融合抗菌肽的植物表達載體,步驟如下:
植物表達載體是由西南大學生物技術中心利用pcambia2300和pbi121自主改造而得的一個雙元表達的基礎載體。該載體的hptii基因用xhoi單酶切后替換為nptii基因(設計引物從pbi121載體中擴增獲得,引物設計兩端帶xhoi位點),限制性酶切和測序結果驗證了nptii基因的方向。在pbi121載體中擴增獲得camv35s啟動子和nos終止子,同時這兩個元件兩端也引入了相應的酶切位點,這兩個元件最后分別導入pcambia的多克隆位點,形成camv35s::mcs::nos單元。該植物表達載體含有1套2×camv35s啟動子控制nptii基因的植物表達元件、1套camv35s啟動子控制報告基因grp-gusplus-his6的植物表達元件和一套camv35s啟動子控制目標基因的植物表達元件,可實現kan和gus活性的雙標記篩選。在多克隆位點(multiplecloningsite,mcs)插入外源基因spcema::erbd,可以實現spcema::erbd的超量表達(圖2)。
煙草遺傳轉化用培養基
共培養培養基(ph5.4):ms無機+b5有機+30g/l蔗糖+2.0mg/l6-ba(6-芐氨基嘌呤)+0.1mg/lnaa(吲哚乙酸)+100μmas(乙酰丁香酮)+2g/lgelrite;
篩選培養基(ph5.8):ms無機+b5有機+30g/l蔗糖+2.0mg/l6-ba(6-芐氨基嘌呤)+0.1mg/lnaa(吲哚乙酸)+400mg/lcef(頭孢噻肟鈉)+100mg/lkm(卡那霉素)+2g/lgelrite;
生根培養基(ph5.8):ms無機+b5有機+30g/l蔗糖+200mg/lcef(頭孢噻肟鈉)+2g/lgelrite。
【實施例3】煙草的遺傳轉化
⑴轉化用農桿菌浸染液的制備
分別挑取含融合抗菌肽基因的根癌農桿菌單菌落,接種于20mlyeb(5g/l蔗糖,1g/l細菌用酵母抽提物,10g/l細菌用胰化蛋白胨,0.5g/lmgso4·7h2o,ph7.0;50mg/lkm(卡那霉素),125mg/lsm(鏈霉素)),28℃、200rpm培養過夜,然后按5%的比例將菌液接種入20ml不含抗生素的液體yeb,28℃、200rpm培養至od600約為0.8-1.2。收集菌液,離心后棄上清,用同等體積的msb液體重懸菌體,然后將菌液全部收集到一個無菌的三角瓶,放在搖床上震蕩培養1h后,即制備成農桿菌浸染液。
⑵煙草遺傳轉化
參考horsch等(science,asimpleandgeneralmethodfortransferringgenesintoplants,1985,227:1229-1231)的方法,以無菌苗的幼嫩葉片為外植體,利用根癌農桿菌介導法進行遺傳轉化。
具體操作為:煙草種子1%次氯酸鈉溶液消毒10min,無菌自來水沖洗5-6次,25℃、16h光照/8h黑暗的光周期于固體萌發培養培養,待無菌苗長至5片葉時,取幼嫩葉片作為農桿菌介導遺傳轉化的外植體。農桿菌浸染液浸染外植體30min后傾去菌液,無菌吸水紙吸去外植體表面多余菌液,然后接種入鋪有一層無菌濾紙的共培養培養基,25℃暗共培養2d。共培養完成后,將外植體接種入篩選培養基中進行分化培養,25℃、16h光照/8h黑暗的光周期培養,每2周繼代一次。產生km抗性幼芽后,將幼芽切下接種入生根培養基,獲得km抗性再生植株。待根長至3-5cm時,對再生植株進行gus染色確認,陽性植株移栽入溫室生長成苗。
gus染色液配方
500mg/lx-gluc(x-gluc溶于dmso,濃度為12.5mg/ml),na2edta(0.01mol/l),k3fe(cn)6(0.1mol/l),k4fe(cn)6(0.1mol/l),tritonx-1001%(v/v),pbs緩沖液(0.14mol/lph7)。
【實施例4】轉基因煙草的gus染色
用打孔器在再生煙草植株的幼嫩葉片上取樣,于37℃恒溫箱中進行gus組織化學染色。3小時后用75%酒精對染色后的樣品進行脫色,并于體視鏡下拍照。結果表明,已成功將目的基因轉入野生型煙草中(圖3)。
【實施例5】轉基因煙草的表達量檢測
為檢測外源基因在轉基因煙草中的表達情況,對溫室中各轉化子植株提取rna,并反轉錄成cdna,利用rt-pcr進行表達量檢測,并基于bio-radcfxmanager和excel進行相應數據的分析。所用引物為(p1:5’-tgcttccttttcttggttc-3’;p2:5’-caacttcaaagctggcaat-3’)。rt-pcr反應體系:5.0μluniversalsybrgreensupermix(bio-rad),4.0μlcdna,0.5μlp1,0.5μlp2,總體積10μl。rt-pcr反應參數:95℃(3min);40個循環:94℃(10sec),56℃(30sec),72℃(30sec)。結果如圖4所示,經rt-pcr的表達量檢測分析,篩選得到了表達量較高的spcema::erbd和spcema轉基因煙草株系各2個。
為了在同一水平上比較spcema:erbd與野生型抗菌肽spcema的轉基因煙草抗病性差異,選取了表達量水平盡可能一致的spcema::erbd-4和spcema-44植株進行抗病鑒定分析。
【實施例6】轉基因煙草葉片離體抗病性比較
首先,檢測了轉基因煙草葉片對大麗輪枝菌的抗性。用不帶針頭的1ml注射器在煙草離體葉片上創建輕微的傷口,隨后移液槍吸取10μl濃度為107個孢子/ml的落葉型大麗輪枝菌的孢子懸浮液注入到傷口處,并在葉柄處包裹上濕潤的吸水紙。將處理后的葉片置于塑料筐中,并覆蓋上一層透明薄膜以保持濕度,于28℃恒溫箱內培養15天后觀察發病情況。結果如圖5a所示,接種病原菌后的野生型煙草葉片發病嚴重,接種處的葉片呈現明顯的褐變現象,并出現大量的真菌菌絲,而接種水的野生型煙草葉片未出現任何病癥,排除了由機械損傷引起葉片病變的可能;野生型spcema轉基因煙草有輕微的病變癥狀,接種處的葉片開始變褐,同時可以觀察到少量的大麗輪枝菌菌絲;而相比野生型spcema,spcema::erbd轉基因煙草葉片在接種處未發現明顯的病變及菌絲。表明融合抗菌肽對大麗輪枝菌具有更強的抗性。
同時,也進行了對煙草赤星病菌的抗性鑒定。取溫室健康生長煙草的第三片葉,利用葉片噴施法接種濃度為107個孢子/ml的煙草赤星病孢子懸浮液,并在葉柄處包裹上濕潤的吸水紙。將處理后的葉片置于塑料筐中,并覆蓋上一層透明薄膜以保持濕度,于28℃恒溫箱內培養15天后觀察發病情況。結果如圖5b所示,接種后的野生型煙草葉片發病嚴重,已呈現明顯的枯萎癥狀;野生型spcema轉基因煙草輕微發病,其葉片的邊緣開始變色、枯萎;而相比野生型spcema,spcema::erbd轉基因煙草葉片上未發現明顯的病斑。表明融合抗菌肽對煙草赤星病菌具有更強的抗性。
【實施例7】轉基因煙草植株總蛋白提取液對病原菌的抗性比較
為了更加直觀的比較spcema::erbd與野生型spcema轉基因煙草的抗性差異,利用0.03mpbs緩沖液(ph5.8)提取植株總蛋白進行體外抑菌實驗。bradford法測定各樣品中總蛋白的含量,并利用蛋白提取液將各植物總蛋白的濃度統一到1mg/ml,隨后比較它們對油菜黑斑病原菌孢子的抑菌效果。
在滅菌處理的90mm培養皿內鋪上一層大小合適的濾紙,加滅菌水濕潤,然后在濾紙上放上兩根牙簽,放上滅菌處理的載玻片。將6μl的煙草蛋白提取液(1mg/ml)、1μl油菜黑斑孢子重懸液(107個/ml)以及3μlpdb滴加在載玻片的中間,于26℃共培養10h后,進行電鏡觀察。結果發現(圖6):油菜黑斑孢子在pbs緩沖液和野生型煙草總蛋白提取液中能夠正常萌發生長;在野生型spcema轉基因煙草總蛋白提取液中,絕大部分的孢子能夠正常萌發,但其菌絲的生長受到了抑制;而融合抗菌肽轉基因煙草總蛋白提取液能夠明顯抑菌孢子的萌發。上述結果表明,麥角甾醇結合域的添加能夠進一步提高spcema對油菜黑斑病原菌孢子的抑菌效果。
sequencelisting
<110>西南大學
<120>一種提高spcema對植物病原菌抗性的方法及轉基因煙草材料
<160>2
<170>patentinversion3.5
<210>1
<211>243
<212>dna
<213>人工序列
<220>
<223>seqidno.1所示的核苷酸序列
<400>1
atggagaagaagtctcttgctggcttgtgcttccttttcttggttctttttgttgctcaa60
gaaattgtggtgactgaagctaagtggaagttgttcaagaagatcggtatcggtgctgtt120
ttgaaggttttgactactggattgccagctttgaagttgaccaagggtggttctggtggt180
ggttctggttttaagttgagagctaagattaaggttagattgagagctaagattaagttg240
tag243
<210>2
<211>80
<212>prt
<213>人工序列
<220>
<223>seqidno.2所示的氨基酸序列
<400>2
mekkslaglcflflvlfvaqeivvteakwklfkkigigavlkvlttglpalkltkggsgg60
gsgfklrakikvrlrakikl80
- 還沒有人留言評論。精彩留言會獲得點贊!