一株耐鹽的木麻黃青枯病生防細菌B268及其應用的制作方法
本發明屬于微生物菌株及其應用,涉及一株一株耐鹽的木麻黃青枯病生防細菌B268,是一株新的耐鹽芽孢桿菌Bacillus siamensis,菌株保藏號為CGMCC No.13225。本發明還涉及該菌株的生防活性,包括抑菌活性和促植物生長活性,及其在木麻黃青枯病防治中的應用。
背景技術:
青枯病又稱細菌性枯萎病(bacterial wilt),是一種由茄科雷爾氏菌(Ralstonia solanacearum)引起的世界范圍內的土傳病害,廣泛發生于熱帶、亞熱帶及溫帶部分地區,近年來由于溫室效應的影響,已出現向高緯度地區蔓延的趨勢。茄科雷爾氏菌種內包含6個生化型(biovar I-VI)和5個生理小種(race 1-5),復雜的種內多樣性使其能夠浸染50多個科、200余種植物,從單子葉到雙子葉,從草本到木本,從灌木到喬木,是寄主范圍最廣的植物病原細菌之一,每年給全球多種經濟作物(如番茄、馬鈴薯、花生、煙草、草莓、香蕉等)和多種林木(如木麻黃、桉樹、桑、油橄欖、柚木、蝴蝶果等)帶來巨大損失,因此被公認為世界第二大最具經濟威脅,同時也最具研究價值的植物病原。
目前,青枯病的防治仍處于研究階段,尚未形成有效手段,而生防微生物是其中最主流的研究方向之一,包括拮抗菌、內生菌和益生菌的使用。綜合文獻查新和專利檢索的結果,迄今已報道的對茄科雷爾氏菌具有生防活性的微生物主要來源于寄主植物的根部土壤和內生環境,包括芽孢桿菌(Bacillus)、短芽孢桿菌(Brevibacillus)、類芽孢桿菌(Paenibacillus)、地芽孢桿菌(Geobacillus)、賴氨酸芽孢桿菌(Lysinibacillus)、假單胞菌(Pseudomonas)、寡養單胞菌(Stenotrophomonas)、不動桿菌屬(Acinetobacter)、腸桿菌屬(Enterobacter)、鏈霉菌(Streptomyces)、節桿菌(Arthrobacter)、弗蘭克氏菌(Frankia)、曲霉(Aspergillus)、木霉(Trichoderma)、青霉(Penicillium)以及噬菌體等,有意思的是,一些分離自海洋環境的芽孢桿菌和鏈霉菌也表現出很高的拮抗活性。芽孢桿菌屬是目前發現青枯病生防菌株最多的一類微生物,有報道的包括解淀粉芽孢桿菌(B.amyloliquefaciens),蘇云金芽孢桿菌(B.thuringiensis),枯草芽孢桿菌(B.subtilis),甲基營養型芽孢桿菌(B.methylotrophicus),死谷芽孢桿菌(B.vallismortis)、蠟狀芽孢桿菌(B.cereus)、巨大芽孢桿菌(‘B.magaterium’)等。而本專利所涉及的新菌株B268與暹羅芽孢桿菌(B.siamensis)的親緣關系最近,雖然已有文獻從基因組角度預測了該種芽孢桿菌的生防潛力,但目前尚未在任何病害的防治研究中有過報道。
另一方面,木麻黃(Casuarina equisetifolia)自引種至我國以來,由于其耐鹽堿、耐潮濕、防風固沙以及速生等特點,已成為我國濱海沙地沿海基干林帶不可替代的先鋒樹種,迄今造林面積已超過30萬公頃。青枯病是木麻黃的主要病害之一,尤其在東南沿海地區,由于氣候溫暖潮濕,有利于茄科雷爾氏菌的生長繁殖,且夏季臺風頻繁,為病菌的侵染和傳播創造了條件,因此,自1964年在廣東省首次發現木麻黃青枯病以來,包括浙江、福建、廣西以及海南等幾乎每個省區均時有發生。茄科雷爾氏菌在野外環境中的潛伏時間長、隱蔽性高,且在木麻黃的苗期到成年期均可引發病害,造成植株大量死亡,嚴重時死亡率可達90%以上,同時還可能交叉浸染發病地區的其他高感病林木以及農作物,危害性大,因此,國家林業局于2013年的第4號公告中將木麻黃青枯病列入全國林業危險性有害生物名單。
迄今,木麻黃青枯病的防治研究以抗病育種技術報道得最多,但考慮到茄科雷爾氏菌的種內變異以及毒性分化,與生物防治技術相結合、‘內外兼修’的綜合防治理念正在成為一種趨勢。例如根據木麻黃的耐鹽耐潮特性,在浙江、福建、廣東一帶有以海水澆淋對木麻黃青枯病進行防治的土法,其基本原理就是通過海水調節植物根部的微生態環境,諸如鹽度、金屬離子濃度、微生物群落結構等,使病原菌的生存和浸染環境惡化,同時有利于特定的生防微生物的生長,具有較好的防治效果。本專利所涉及的耐鹽芽孢桿菌Bacillus siamensis B268即分離自海水浸漬后的木麻黃根部土壤,該菌株兼具病原菌拮抗活性以及促植物生長活性,且對海鹽的耐受范圍廣,定殖能力強,在木麻黃青枯病的防治中具有一定的應用開發潛力,同時,在我國土地鹽漬化情況日趨嚴峻的大背景下,該菌株的研究對于各種土質下的植物病害防治也具有一定的借鑒意義。
技術實現要素:
本發明的一個目的是提供一株耐鹽的木麻黃青枯病生防細菌B268,是一株新的芽孢桿菌。本發明涉及的菌株B268來源于感病木麻黃植株,采集海水浸漬后的根部土壤,通過對峙板活性篩選獲得。進一步的分析結果表明,該菌株歸屬于暹羅芽孢桿菌(Bacillus siamensis),且顯示出較強的生防活性,包括:(1)對分離自同一木麻黃林區的茄科雷爾氏菌致病株Ralstonia solanacearum YQ以及購買自比利時微生物菌種保藏中心(Belgian Coordinated Collections of Microorganisms,BCCM)的茄科雷爾氏菌標準株Ralstonia solanacearum LMG 2299T均具有拮抗活性;(2)具有解磷、生成嗜鐵素以及吲哚乙酸等促植物生長活性;(3)海鹽的耐受范圍廣,而且能在不同的培養基以及多個鹽度下形成生物被膜,顯示出較強的定殖能力。目前,Bacillus siamensis菌株在青枯病防治的研究和應用中尚未見報道。所述的菌株B268已被中國微生物菌種保藏管理委員會普通微生物中心保藏(地址:北京市朝陽區北辰西路1號院3號,中國科學院微生物研究所,郵編100101),分類命名為Bacillus siamensis,保藏號為CGMCC No.13225,保藏日期為2016年11月1日。
本發明的另一個目的是提供所述菌株B268在木麻黃青枯病的生物防治中的用途。所述的菌株Bacillus siamensis B268可用于制備木麻黃青枯病的生物防治菌劑或農藥,或者用于制備促木麻黃生長及提高其抗病性的微生物菌肥。所述的菌株Bacillus siamensis B268用于制備促木麻黃生長及提高其抗病性的微生物菌肥。實驗證明所述的菌株Bacillus siamensis B268,其對于木麻黃及番茄青枯病的的致病菌株均具有拮抗活性。所述的菌株Bacillus siamensis B268,其具有促植物生長活性,包括解磷活性,合成并分泌嗜鐵素的活性,以及合成植物生長激素——吲哚乙酸(IAA)的活性。所述的菌株Bacillus siamensis B268,其分離自海水浸漬的木麻黃根部土壤,具有寬的海鹽耐受范圍(0-12%),以及在有鹽和無鹽條件下(0-5%)生成穩定生物膜的能力。
目前,涉及青枯病防治的菌株以煙草青枯病居多,其他還包括番茄、花生、馬鈴薯、辣椒、生姜、茄子等作物青枯病,而木麻黃青枯病則尚未有報道。
為實現上述目的,本發明采取的技術方案為:
(1)菌株Bacillus siamensis B268的分離及拮抗活性篩選。據文獻報道,海洋環境中存在著不少對茄科雷爾氏菌具有高拮抗活性的微生物,同時,海水澆淋法能夠有效抑制發病初期的木麻黃青枯病,結合以上信息,專門聯系了浙江省樂清市森林病蟲防治檢疫站開展合作,在其對海水澆淋法進行防治效果驗證的木麻黃林區內采集根部土壤進行微生物的分離。同時采集林區中重病木麻黃植株的木質部樣品,進行病原菌株的分離。最后通過對峙板法篩選獲得多株拮抗微生物,其中即包括菌株B268。
(2)菌株Bacillus siamensis B268的拮抗活性驗證。在已獲得的木麻黃青枯病致病株Ralstonia solanacearum YQ的基礎上,從比利時微生物菌種保藏中心(Belgian Coordinated Collections of Microorganisms,BCCM)購買青枯病原菌的標準株Ralstonia solanacearum LMG 2299T,兩株菌分屬于茄科雷爾氏菌的種內不同生化型,后者分離自番茄(Lycopersicon esculentum)青枯病病株,同時將兩者作為靶標對菌株B268的拮抗活性進行驗證(圖3)。
(3)促植物生長活性分析。研究表明,解磷菌能夠將土壤中植物難以吸收利用的不溶性磷轉化為可吸收利用的可溶性磷,同時,一些微生物能夠分泌小分子的鐵螫合劑——嗜鐵素(Siderophore),通過對三價鐵的特異性結合幫助植物從土壤中富集這種限制性營養元素,這些生理特性有利于改善植物根部的營養環境,從而間接地促進植物生長。此外,根據文獻報道,很多生防菌都能夠合成植物內源性的生長激素——吲哚乙酸(indole-3-acetic acid,
IAA),通過直接調節植物生長來抵御病原菌的侵襲。基于上述信息,分別對菌株B268的解磷活性、吲哚乙酸生成以及嗜鐵素生成進行了檢測(圖4-6)。
(4)鹽的耐受性分析。鑒于菌株B268分離自海水浸漬后的根土,金屬元素分析結果表明,土樣中的鈉鎂離子濃度均明顯高于普通土壤。同時,與B268親緣關系最近的菌株Bacillus siamensis PD-A10T分離自鹽漬螃蟹,能夠耐受14%(w/v)的NaCl濃度。基于上述背景,對菌株B268的耐鹽性進行了分析驗證(圖7)。
(5)生物被膜的形成能力分析。生物被膜(biofilm)是細菌適應環境的一種特殊生長狀態,即面對環境脅迫時的一種自我調控方式,與細菌的定殖、浸染、毒力等多種特性密切相關,而定殖能力又是決定生防菌株能否在林間長期發揮防治效用,進而決定其是否具有實際應用價值的重要指標之一,基于上述原因,對菌株B268的生物被膜形成能力進行了考察(圖8)。
(6)木麻黃青枯病的防治活性驗證。以溫室培養的盆栽木麻黃苗為寄主,Ralstonia solanacearum YQ為病原,對菌株B268的生防活性進行了比較和驗證(圖9)。
綜上所述,本發明提供的菌株Bacillus siamensis B268豐富了生防芽孢桿菌的種類及其施用范疇,該菌株在不同培養基中均表現出強的生長能力及穩定的活性,具有進一步研發的價值。
本發明提供一株耐鹽芽孢桿菌B268,該菌株分離自海水浸漬的木麻黃根部土壤,對木麻黃及番茄青枯病的致病菌株均具有拮抗活性,同時具有解磷、合成嗜鐵素以及植物生長激素等促植物生長活性。此外,該菌株對海鹽的耐受范圍廣,能在多個鹽度下形成穩定的生物膜,生長及定殖能力強。鑒于上述生防活性,本發明提供的菌株B268可在制備木麻黃青枯病的生物防治中的應用,基于該菌株B268對病原菌的拮抗活性以及強的生物膜形成能力,能開發為病害防治菌劑或直接生產為生物農藥,基于解磷、分泌嗜鐵素以及IAA的活性,菌株B268能開發為菌肥。
附圖說明
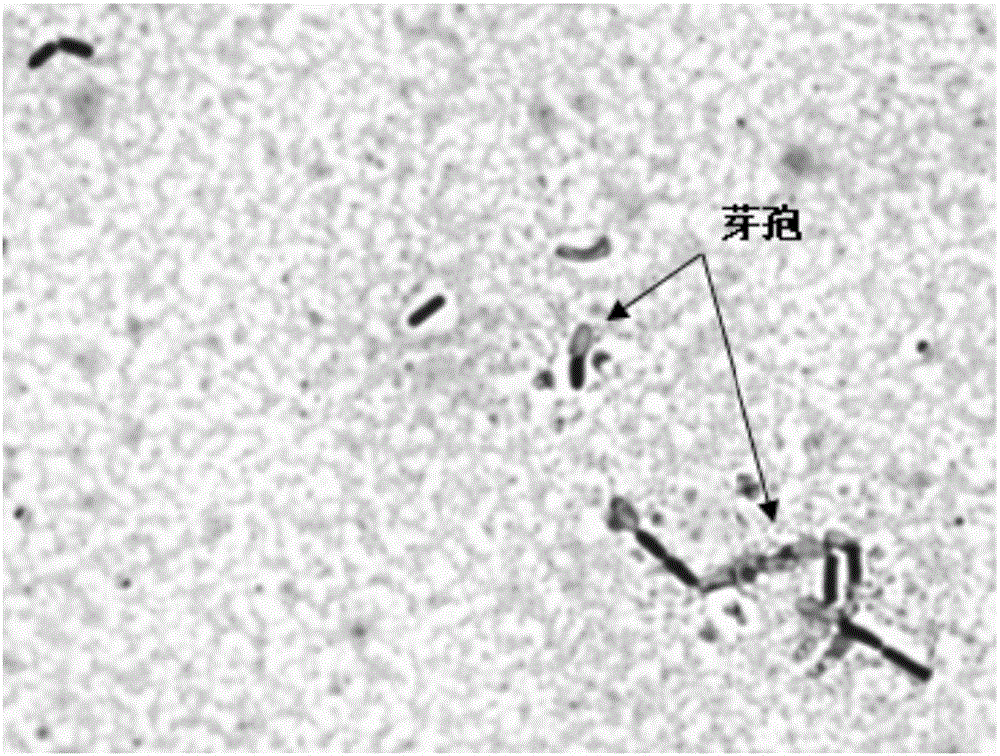
圖1菌株Bacillus siamensis B268的細胞形態。
圖2菌株Bacillus siamensis B268及其近緣菌株的Neighbor-Joining系統發育樹。
圖3菌株Bacillus siamensis B268對茄科雷爾氏菌的拮抗活性,其中A:富營養人工海水培養基+病原菌株Ralstonia solanacearum YQ+B268(活性篩選),B1:富營養人工海水培養基+病原菌株Ralstonia solanacearum YQ+B268(活性驗證),B2:富營養人工海水培養基+病原菌株Ralstonia solanacearum LMG 2299T+B268,B3:Modified YPG無鹽培養基+病原菌株Ralstonia solanacearum YQ+B268,B4:Modified YPG無鹽培養基+病原菌株Ralstonia solanacearum LMG 2299T+B268。
圖4菌株Bacillus siamensis B268的解磷活性,其中4-1:Pikovskaya Agar平板,4-2:NBRI-BPB平板。
圖5菌株Bacillus siamensis B268的嗜鐵素生成。
圖6菌株Bacillus siamensis B268的IAA生成,其中A:Modified YPG無鹽培養基檢測結果,B:2%海鹽+Modified YPG無鹽培養基檢測結果;對照1:補加色氨酸但不接種培養管,平行1-3:補加色氨酸并接種培養管,對照2:不補加色氨酸但接種培養管。
圖7菌株Bacillus siamensis B268的耐鹽性。
圖8A菌株Bacillus siamensis B268在不同鹽度下的生物被膜形成的結晶紫洗脫液的顏色對比。
圖8B菌株Bacillus siamensis B268在不同鹽度下的生物被膜形成的結晶紫洗脫液的OD570檢測結果。
圖9菌株Bacillus siamensis B268對木麻黃青枯病的防治效果,其中9-1:不同菌液灌根處理后的木麻黃苗的根部外觀,9-2:不同菌液灌根處理后的木麻黃苗的莖基部橫切面;圖中A:無菌培養基+病原菌株Ralstonia solanacearum YQ培養液處理;B:Bacillus subtilis NH2-51-2培養液+病原菌株Ralstonia solanacearum YQ培養液處理;C:Bacillus siamensis B268培養液+病原菌株Ralstonia solanacearum YQ培養液處理。
具體實施方式
結合附圖和實施例對本發明作進一步的說明。
實施例1:菌株Bacillus siamensis B268的分離及活性篩選
樣品采集自浙江省樂清市濱海木麻黃人工林,該林區曾進行過海水澆淋對木麻黃青枯病防治效果的驗證性實驗,在海水浸漬過的林區中,根據目測的發病狀況分別選取無病癥和有病癥植株,采集根圍10cm以下的土壤。
在菌株分離前,對樣品鹽度進行驗證,即采用電感耦合等離子體發射光譜法(Inductively Coupled Plasma Optical Emission Spectrometry,ICP-OES)對土樣的金屬元素組成及含量進行分析,結果表明,其中的鈉鎂離子濃度分別為0.5-0.7%和0.3-0.5%,明顯高于作為對照的普通土樣。
取10g土樣以20ml無菌生理鹽水懸浮,于28℃、140rpm條件下浸提2天。懸浮液以無菌生理鹽水梯度稀釋后涂布寡營養人工海水平板,于28℃培養至形成明顯菌落。根據形態差異分別挑取單菌落,以相同平板進行菌株的純化,并以20%(v/v)甘油管形式保藏。
分離菌株以富營養人工海水平板劃線活化,28℃培養24小時,病原菌株Ralstonia solanacearum YQ以富營養人工海水液體搖管活化,28℃、140rpm培養48小時。取50μl菌株YQ的培養液(OD600=1.0)劃線富營養人工海水平板,再挑取3個分離菌株的單菌落點種該平板,制成的對峙板于28℃培養3天,逐日觀察抑菌圈的形成情況。菌株B268的活性篩選結果見圖3A。
人工海水:NaCl 4.86g,MgCl2·6H2O 3.15g,MgSO4·7H2O 1.66g,CaCl2 0.45g,KCl 0.138g,NaHCO3 40mg,檸檬酸鐵2.5mg,SrCl2·6H2O 14.25mg,KBr 20mg,H3BO3 5.5mg,Na2SiO3·9H2O 2.32mg,NaF 0.6mg,NH4NO3 0.6mg,Na2HPO4 2mg,去離子水1L。
寡營養人工海水培養基:Yeast Extract(BD 212750)0.5g,Trypticase Peptone(BD 211921)0.5g,人工海水1L,pH 7.0。
富營養人工海水培養基:Yeast Extract(BD 212750)1g,Trypticase Peptone(BD 211921)5g,人工海水1L,pH 7.0。
目前,菌株B268已在中國微生物菌種保藏管理委員會普通微生物中心保藏(地址:北京市朝陽區北辰西路1號院3號,中國科學院微生物研究所,郵編100101),分類命名為Bacillus siamensis,保藏號為CGMCC No.13225,保藏日期為2016年11月1日。
實施例2:菌株Bacillus siamensis B268的系統發育鑒定
挑取菌株B268的平板單菌落,轉接富營養人工海水培養基,28℃、140rpm培養24小時后收集細胞,鏡檢的同時采用細菌基因組快速抽提試劑盒(東勝生物)抽提基因組模板。采用細菌通用引物27F(5'-AGAGTTTGATCCTGGCTCAG-3')以及1492R(5'-ACGGTTACCTTGTTACGACTT-3')擴增16S rRNA基因,獲得的產物經AxyPrep PCR清潔試劑盒(Axygen)純化后與pMD 19-T Vector連接(TaKaRa),并轉化Trans1-T1感受態細胞(TransGen Biotech),挑取陽性重組子送交測序公司進行測序。
顯微鏡下菌株B268成桿狀,可觀察到芽孢的形成(圖1)。
菌株B268的16S rRNA基因全長1513nt(詳見序列表),序列已遞交至GenBank數據庫,序列登錄號為KY129717。采用EzBioCloud(http://www.ezbiocloud.net/)比對工具進行兩兩序列間的相似性分析,結果表明,B268與Bacillus siamensis KCTC 13613T的序列相似性最高,為99.93%。采用Mega 6.0進行多序列比對和系統發育關系分析,結果表明,菌株B268在Neighbor-Joining系統發育樹上與芽孢桿菌屬的標準種B.subtilis,以及經典的青枯病生防菌B.amyloliquefaciens,B.methylotrophicus,潛在的生防菌B.siamensis處于同一個進化分支,且與Bacillus siamensis KCTC 13613T間的進化距離最短(圖2)。
綜合上述分析結果,將菌株B268劃分至芽孢桿菌屬中的Bacillus siamensis種。
菌株Bacillus siamensis B268保藏于中國微生物菌種保藏管理委員會普通微生物中心,保藏號為CGMCC No.13225。
實施例3:菌株Bacillus siamensis B268的拮抗活性驗證
分別使用富營養人工海水培養基及Modified YPG無鹽培養基制備對峙板,分別以本實驗室分離自木麻黃病株的Ralstonia solanacearum YQ以及購買自BCCM的分離自番茄病株的Ralstonia solanacearum LMG 2299T為靶標進行拮抗活性的驗證。菌株B268以平板劃線活化,28℃培養24小時,兩株病原菌株以液體搖管活化,使用富營養人工海水培養基時,于28℃、140rpm培養48小時,使用Modified YPG無鹽培養基時,于28℃、140rpm培養24小時。對峙板的制備方法同實施例1,于28℃培養3天后觀察抑菌圈的形成情況。
如圖3B所示,病原菌株YQ及LMG 2299T在無鹽培養基上生長更好,而菌株B268在兩種培養基上均生長良好,且對兩株侵染不同寄主的病原菌均表現出明顯的拮抗活性。
Modified YPG無鹽培養基:牛肉浸粉3g,蛋白胨5g,葡萄糖10g,酵母抽提物0.5g,蒸餾水1L,pH 7.0;
實施例4:菌株Bacillus siamensis B268的解磷試驗
菌株B268劃線富營養人工海水平板活化,28℃培養24小時后點種Pikovskaya Agar平板以及NBRI-BPB平板以鑒定其解磷活性,28℃培養至出現明顯解磷圈。結果如圖4-1及4-2所示,Pikovskaya Agar平板上出現透明圈而NBRI-BPB平板上出現黃色圈,表明菌株B268具有解磷活性。
Pikovskaya Agar培養基:Yeast Extract(BD 212750)0.5g,葡萄糖10g,Ca3(PO4)25g,(NH4)2SO4 0.5g,K2SO4 0.2g,MgSO4·7H2O 0.1g,MnSO4 2mg,FeSO4·7H2O 2mg,瓊脂粉15g,蒸餾水1L,pH 7.0;
NBRI-BPB培養基:葡萄糖10g,Ca3(PO4)2 5g,MgCl2·6H2O 5g,MgSO4·7H2O 0.25g,KCl 0.2g,(NH4)2SO4 0.1g,溴酚藍25mg,瓊脂粉15g,蒸餾水1L,pH 7.0;
實施例5:菌株Bacillus siamensis B268的嗜鐵素合成試驗
菌株B268劃線富營養人工海水平板活化,28℃培養24小時后點種LBA平板以鑒定其嗜鐵素合成活性,28℃培養至出現明顯反應圈。結果如圖5所示,平板上出現黃色透明圈,表明菌株B268能夠合成并分泌嗜鐵素。
LBA培養基:60.5mg鉻天青S溶于50ml去離子水中,以10mM HCl配制1mM的FeCl3·6H2O溶液,取該鐵溶液10ml加入鉻天青S水溶液中混勻,形成CAS溶液;72.9mg CTAB溶于40ml去離子水中,邊攪拌邊緩慢加入上述CAS溶液,混勻后滅菌備用;
Tryptone(BD 211705)10g,Yeast Extract(BD 212750)5g,NaCl 10g,PIPES 30.2g,去離子水900mL,以50%NaOH溶液調pH 6.8,加瓊脂粉15g后滅菌備用;
上述兩種基質混合后倒制LBA平板;
實施例6:菌株Bacillus siamensis B268的IAA合成試驗
菌株B268以Modified YPG無鹽培養基活化,轉接相同培養基后補充終濃度為40mg/l的色氨酸,28℃、140rpm培養,以接菌但不補加色氨酸以及補加色氨酸但不接菌的平行培養管為對照。同時轉接補加2%海鹽(Sigma S9883)的Modified YPG無鹽培養基,其余操作相同。
取7天培養液1ml,與等體積檢測試劑混合后,室溫避光放置24小時,期間觀察顏色變化。結果如圖6A所示,凡接種的培養管,無論是否補加色氨酸,加入檢測試劑后均顯紅色,表明菌株B268能夠合成IAA。而圖6中B的結果與A相近,表明鹽對于菌株B268的IAA合成無明顯影響。
檢測試劑:12g FeCl3·6H2O,7.9M硫酸1L;
實施例7:菌株Bacillus siamensis B268的耐鹽性分析
在Modified YPG無鹽培養基的基礎之上添加0-12%海鹽(Sigma S9883),配制鹽梯度培養基,分別接種菌株B268以及病原菌株YQ和LMG 2299T,28℃、140rpm培養24小時后檢測OD600值。結果如圖7所示,菌株B268對海鹽的耐受范圍較廣,培養24小時時,在1-7%的海鹽濃度下均生長良好(OD600>1),而延長培養時間時,在0-12%的海鹽濃度下均生長良好(OD600>1)。相反的,病原菌株YQ和LMG 2299T均不耐鹽,明顯喜好無鹽或低鹽環境(<1%),當海鹽濃度大于3%時即不能生長。而根據文獻報道,木麻黃的鹽耐受范圍為0.3-1.5%,與病原菌相當。基于上述數據可以認為:一,菌株B268的耐鹽性與其分離自海水浸漬的土壤有關;二,寬的海鹽耐受范圍使得菌株B268易于在內陸、濱海甚至灘涂等不同含鹽量的木麻黃種植帶土壤中生存;三,耐鹽性的差異預示了菌株B268可能協同含鹽環境共同拮抗病原菌,而這可能是海水澆淋法抑制木麻黃青枯病的機制之一。
實施例8:菌株Bacillus siamensis B268的生物膜形成試驗
在Modified YPG無鹽培養基的基礎之上添加不同濃度的海鹽(Sigma S9883),制備鹽梯度培養基,以195μl/孔的加量將上述海鹽梯度培養基分別加入96孔培養板,每個梯度6個平行孔。以Modified YPG無鹽培養基制備病原菌株YQ,LMG 2299T以及菌株B268的種子液,調OD600=0.3后,按5μl/孔的加量接種,同時以加入5μl無菌培養基的孔作為陰性對照。培養板的具體布局如表1所示,28℃靜置培養20小時后進行生物膜的檢測,具體方法如下:
以微量移液器小心吸走每孔中的菌液以及液體表面形成的菌膜,以無菌水輕柔洗滌每孔兩次;吸干殘余液體后按300μl/孔的加量加入1%(w/v)結晶紫水溶液,室溫下靜置染色30分鐘;小心吸走染液,在合適的容器中以無菌水輕柔漂洗孔板至無明顯浮色為止;吸干殘余液體后按300μl/孔的加量加入33%(v/v)乙酸溶液,室溫下靜置,其目的是洗脫被生物膜固定的結晶紫,充分洗脫后檢測OD570值。
表1
如圖8A及圖8B所示,菌株B268在0-5%的海鹽濃度下均能形成穩定的生物膜,尤以1-3%的海鹽濃度為最適(洗脫結晶紫的顏色最深或OD570值最高),這一趨勢與其耐鹽性基本一致,說明適當濃度的海鹽能夠促進菌株B268的定殖。而病原菌株YQ僅在0-1%海鹽濃度下形成微弱的生物膜,LMG 2299T僅在0.5%海鹽濃度下形成生物膜,表明在相同的營養條件下,菌株B268更具有生長優勢,若在野外環境下,則意味著生態位的優先占據,從而達到抑制病原菌增殖的目的。
實施例9:菌株Bacillus siamensis B268對木麻黃青枯病的防治試驗
菌株B268、Bacillus subtilis NH2-51-2及病原菌株YQ分別接種Modified YPG無鹽培養基,28℃、140rpm培養24小時。菌液調OD600=1后,分別將NH2-51-2與YQ,B268與YQ,無菌培養基與YQ等體積混合。其中,菌株NH2-51-2分離自南海表層海水,對峙板篩選結果顯示其對病原菌株YQ有微弱的拮抗活性,在本實驗中作為陽性對比菌株。加入無菌培養基的YQ菌液作為陰性對照。以灌根法分別將上述3種混合菌液以5ml/株接種盆栽木麻黃苗,溫室培養1個月后取根至莖基部分鑒定侵染情況。
如圖9所示,在相同的培養時間下,僅接種病原菌株YQ的植株,根表呈黑色(圖9-1A),莖基橫切面干枯且維管組織有明顯的腐爛現象(圖9-2A),出現了青枯病侵染的癥狀;接種菌株B268及YQ混合液的植株,根表呈健康的亮褐色,且有光澤(圖9-1C),莖基橫切面新鮮濕潤(圖9-2C),未出現青枯病侵染的癥狀;而接種菌株NH2-51-2及YQ混合液的植株,根表呈暗褐色(圖9-1B),莖基橫切面略顯干枯,維管組織有輕微腐爛痕跡(圖9-2B)。上述結果表明,菌株NH2-51-2拮抗活性較弱,能延緩但不能抑制病害的發生,而菌株B268的存在能有效抑制病原菌對木麻黃根部的侵入,具有病害防治的效果。
<110> 浙江農林大學
<120> 一株耐鹽的木麻黃青枯病生防細菌B268及其應用
<160> 3
<210> 1
<211> 1513
<212> DNA
<213> Bacillus siamensis
<400> 1
1 AGAGTTTGAT CCTGGCTCAG GACGAACGCT GGCGGCGTGC CTAATACATG CAAGTCGAGC
61 GGACAGATGG GAGCTTGCTC CCTGATGTTA GCGGCGGACG GGTGAGTAAC ACGTGGGTAA
121 CCTGCCTGTA AGACTGGGAT AACTCCGGGA AACCGGGGCT AATACCGGAT GGTTGTTTGA
181 ACCGCATGGT TCAGACATAA AAGGTGGCTT CGGCTACCAC TTACAGATGG ACCCGCGGCG
241 CATTAGCTAG TTGGTGAGGT AACGGCTCAC CAAGGCGACG ATGCGTAGCC GACCTGAGAG
301 GGTGATCGGC CACACTGGGA CTGAGACACG GCCCAGACTC CTACGGGAGG CAGCAGTAGG
361 GAATCTTCCG CAATGGACGA AAGTCTGACG GAGCAACGCC GCGTGAGTGA TGAAGGTTTT
421 CGGATCGTAA AGCTCTGTTG TTAGGGAAGA ACAAGTGCCG TTCAAATAGG GCGGCACCTT
481 GACGGTACCT AACCAGAAAG CCACGGCTAA CTACGTGCCA GCAGCCGCGG TAATACGTAG
541 GTGGCAAGCG TTGTCCGGAA TTATTGGGCG TAAAGGGCTC GCAGGCGGTT TCTTAAGTCT
601 GATGTGAAAG CCCCCGGCTC AACCGGGGAG GGTCATTGGA AACTGGGGAA CTTGAGTGCA
661 GAAGAGGAGA GTGGAATTCC ACGTGTAGCG GTGAAATGCG TAGAGATGTG GAGGAACACC
721 AGTGGCGAAG GCGACTCTCT GGTCTGTAAC TGACGCTGAG GAGCGAAAGC GTGGGGAGCG
781 AACAGGATTA GATACCCTGG TAGTCCACGC CGTAAACGAT GAGTGCTAAG TGTTAGGGGG
841 TTTCCGCCCC TTAGTGCTGC AGCTAACGCA TTAAGCACTC CGCCTGGGGA GTACGGTCGC
901 AAGACTGAAA CTCAAAGGAA TTGACGGGGG CCCGCACAAG CGGTGGAGCA TGTGGTTTAA
961 TTCGAAGCAA CGCGAAGAAC CTTACCAGGT CTTGACATCC TCTGACAATC CTAGAGATAG
1021 GACGTCCCCT TCGGGGGCAG AGTGACAGGT GGTGCATGGT TGTCGTCAGC TCGTGTCGTG
1081 AGATGTTGGG TTAAGTCCCG CAACGAGCGC AACCCTTGAT CTTAGTTGCC AGCATTCAGT
1141 TGGGCACTCT AAGGTGACTG CCGGTGACAA ACCGGAGGAA GGTGGGGATG ACGTCAAATC
1201 ATCATGCCCC TTATGACCTG GGCTACACAC GTGCTACAAT GGACAGAACA AAGGGCAGCG
1261 AAACCGCGAG GTTAAGCCAA TCCCACAAAT CTGTTCTCAG TTCGGATCGC AGTCTGCAAC
1321 TCGACTGCGT GAAGCTGGAA TCGCTAGTAA TCGCGGATCA GCATGCCGCG GTGAATACGT
1381 TCCCGGGCCT TGTACACACC GCCCGTCACA CCACGAGAGT TTGTAACACC CGAAGTCGGT
1441 GAGGTAACCT TTATGGAGCC AGCCGCCGAA GGTGGGACAG ATGATTGGGG TGAAGTCGTA
1501 ACAAGGTAGC CGT
<210> 2
<211> 20
<212> DNA
<213> 人工序列
<223> 16S rRNA基因上游引物
<400> 2
AGAGTTTGAT CCTGGCTCAG
<210> 3
<211> 21
<212> DNA
<213> 人工序列
<223> 16S rRNA基因下游引物
<400> 3
ACGGTTACCT TGTTACGACT T
- 還沒有人留言評論。精彩留言會獲得點贊!