對綠盲蝽進行RNA干擾的飼喂方法及其在基因篩選上的應用與流程
本發明涉及生物應用技術領域,特別是涉及對綠盲蝽進行RNAi的飼喂方法以及利用該方法在篩選綠盲蝽生長發育關鍵基因上的應用。
背景技術:
1997年開始的Bt棉花種植規模的擴大導致以前處于次要害蟲的綠盲蝽代替棉鈴蟲成為對我國農業生產嚴重為害的主要害蟲,特別是成為長江和黃河流域棉田的重要害蟲,在此地區綠盲蝽發生面積甚至達到了90%左右,不但造成棉鈴掉落而且葉子枯萎從而嚴重影響棉花產量,所產生的經濟損失產量損失約10-20%,嚴重發生地區超過30%。此外,綠盲蝽不但危害棉花,而且還影響蘋果、葡萄、櫻桃、栆等其他水果和農作物的產量和質量。當下防治此害蟲的方法很多,比如生物防治,昆蟲綜合防治,化學防治和物理防治等。遺憾的是至今綠盲蝽還沒有一種專一性天敵,而且防治成本很高,在Bt蛋白方面還沒有找到效果很好的一種防治蛋白,所以綠盲蝽的防治被迫靠化學方法來進行。雖然化學農藥的使用有嚴格的執行規律,但反復使用農藥,不但嚴重污染環境,而且綠盲蝽對一些農藥也開始產生抗藥性,而且許多殺蟲劑嚴重的缺點是也可以殺死非靶標的有益昆蟲。鑒于將來發展趨勢,開發一條田間可行,經濟效益高、環境友好型地綠色防治途徑是擺在研究人員面前的重要任務。
雙鏈RNA(dsRNA)介導的基因沉默通常被稱為RNA干擾(RNAi),當外源雙鏈dsRNA(長雙鏈RNA)或siRNA(小干擾RNA)被轉入到細胞或組織時在體內產生內源互補靶標mRNA序列的特異性降解并阻斷轉錄,抑制翻譯的現象。在這個過程中首先dsRNA被Dicer酶切割成21-23bp長度的小分子RNA即siRNA,然后RNA誘導沉默復合體(RNA-induced silencing complex,RISC)的作用下siRNA被解旋成單鏈,反義鏈特異性地與同源mRNA(靶標)結合并引導RISC使mRNA發生斷裂而導致基因表達沉默。RNAi的發現在生物領域徹底改變了研究方法和思考模式。通過將近20年的研究RNAi已變成了非常成熟的反向遺傳學研究基因功能的巧妙工具。至今RNAi在醫學,作物改良,農業,畜牧業,園藝,農業等方面取得了很大的進展。理論上講我們通過RNAi可也沉默任何一個我們感興趣的靶標基因。至今研究人員在這些方面做了大量研究篩選出大量的靶標基因,并實際應用取得了令人興奮地突破。比如Baum(Baum,J.A.et al.(2007)Control of coleopteran insect pests throughRNA interference.Nat.Biotechnol.25,1322–1326)等通過飼喂方法在西方玉米根蟲(WCR)中一共篩選將近300中基因并最后證明14種基因的RNAi效果是很好的。
自從1998年首次在昆蟲里被發現以來,在幾乎所有昆蟲種類都發現RNAi現象并初步進入應用階段。最近通過細菌、寄主植物和寄主植物病毒成功的用于干擾昆蟲靶標基因的表達。這些研究使用了轉基因介導的RNAi并在害蟲防治方面取得了顯著地進步。在細菌里靶標昆蟲的dsRNA通過把部分cDNA在T7啟動子中反向定位來構建載體在桿菌球菌Escherichia coli Migula菌株中表達而產生的,這樣dsRNA在細菌細胞里像體外合成那樣產生。吸收這樣表達dsRNA的細菌顯示在草地夜蛾Spodoptera frugiperda、橘小實蠅Bactrocera dorsalis Hendel和馬鈴薯甲蟲Leptinotarsadecemlineata Say不但在表達量而且表型上產生強大的RNAi反應。這些dsRNA表達細菌相當于新的生物農藥。然而為提高效率和田間使用需要新的傳遞方法,因此發展能夠誘導RNAi的轉基因植物最近幾年引起了很大關注。在這個系統里寄主植物被帶有靶標昆蟲基因反向重復序達載體的土壤農桿菌侵染并形成能夠表達dsRNA的轉基因植物。至今這種方法分別在玉米根螢葉甲virgifera LeConte,棉鈴蟲Helicoverpa armigera Hübner(Mao et al.,2007),煙草天蛾天蛾M.sexta和一些吸韌皮部汁液昆蟲如褐飛虱Nilaparvata lugens 桃蚜Myzus persicae Sulzer等中產生明顯的死亡率和昆蟲蛻皮異常等表型。這些研究成果使RNAi在田間應用方面邁出了突破性的一步。至今昆蟲里dsRNA的傳遞方法主要有顯微注射法,飼喂即口服傳遞法,轉基因植物法等三種。由于利用RNAi技術培育轉基因植物,周期長,成本高,不能實現從大量基因中篩選高效率基因,所以在發展轉基因植物之前能夠尋找既經濟又簡單快捷的方法篩選鑒定大量基因從而找到候選基因顯得尤為重要。顯微注射方法最致命的缺點是在實驗昆蟲里產生機械性死亡而影響死亡結果,此外考慮到將來田間應用dsRNA,必須通過昆蟲消化道并在這樣特別不友好的環境里持續存在一段時間,所以在實驗室初步篩選階段,最有說服力的dsRNA傳遞方法無疑是飼喂即口服方法。飼喂法相對于注射法,有實驗成本低,便于大規模實驗,不易造成昆蟲人為原因而死亡等顯著優勢。雖然前一段時間關于綠盲蝽RNAi的研究報較多,然而1)主要集中在嗅覺系統相關基因上,RNAi介導的殺蟲劑方面的研究處于空白狀態,2)上述研究都使用顯微注射dsRNA傳遞方法。所以發展以飼喂RNAi介導的dsRNA傳遞方法是大大促進基因篩選和將來田間應用的腳步。
Actin即肌動蛋白是廣泛存在于細胞的一種重要骨架蛋白,Actin在細胞吞噬、分泌、移動、胞質流動和胞質分離等過程中起重要作用。其中β-Actin是橫紋肌肌纖維中的一種主要蛋白質成分,也是肌肉細絲及細胞骨架微絲的主要成分,具有收縮功能,分布廣泛。因為其在細胞里非常保守及表達量相對恒定,所以β-Actin一般作為內參基因來檢測。所以我們首先用β-Actin來驗證了綠盲蝽飼喂RNAi存在與否。
V-ATPase是廣泛存在于原核生物和真核細胞的胞質膜和管泡膜膜系統中的持家基因,其由8個亞基(A、B、C、D、E、F、G、H)構成的V1催化結構域和6個亞基(a、c、c′、c″、d、e)構成的膜融合V0結構域組成。V-ATPaseE是主要亞基之一。
技術實現要素:
本發明提供一種對綠盲蝽進行RNAi實驗的飼喂方法,首先在證明綠盲蝽飼喂RNAi存在的基礎上對綠盲蝽進行RNAi實驗的飼喂參數(包括喂食濃度、喂食體積,喂食dsRNA總量)進行確定,并挑選了綠盲蝽細胞膜及管泡細胞器膜上廣泛分布一種與H+主動轉運有關的蛋白E亞基即V-ATPaseE作為靶標基因驗證了綠盲蝽口服傳遞的dsRNA的RNAi效果,結果表明導入V-ATPaseE基因dsRNA可以在綠盲蝽體內誘導RNAi反應并引起靶標害蟲快速死亡。該方法為篩選可靠,高效率,可持續的抗蟲基因提供新的技術手段和平臺,為將來發展抗綠盲蝽轉基因棉花的培育打下了踏實的基礎。
綠盲蝽飼喂RNAi實驗的方法,是把靶標基因dsRNA混合到綠盲蝽人工飼料中,并通過綠盲蝽吸入體內來進行RNA干擾。
人工飼料中混合dsRNA最終濃度為0.3-0.8μg/μl,每天喂食體積為10-15μl/頭,所述持續飼養時間為5-7天。
優選人工飼料中混合dsRNA最終濃度為0.5μg/μl,每天喂食體積200μl/15頭。
所述人工飼料包裹在封口膜內。
所述綠盲蝽為3L若蟲。
上述方法在綠盲蝽生長發育關鍵基因的篩選中的應用。
所述應用包括如下步驟:
(1)測序綠盲蝽轉錄組并建立本地BLAST數據庫,通過NCBI已公布的其他物種靶標基因序與轉錄組測序所獲得的比對篩選靶標基因并確定基因片段,(2)取綠盲蝽總RNA,通過RT-PCR得到cDNA;(3)設計帶有T7序列的特異性引物;(4)通過PCR技術擴增目的基因片段:以cDNA為模板,以設計的特異性引物擴增,得到帶有T7序列和目的基因片段的PCR產物;(5)構建含有選定目的基因片段的載體并轉入宿主大腸桿菌中,培養,提取含有目的基因片段的質粒;(6)以該質粒為模板,以設計的帶有T7序列的特異性引物擴增,得到大量含有T7序列和目的基因片段的PCR產物;(7)利用T7RNA聚合酶將上一步得到的PCR產物合成為目的基因片段dsRNA;(8)將dsRNA混合到人工飼料中并飼喂綠盲蝽3齡幼蟲24小時后用正常飼料繼續飼養;(9)統計死亡率,觀察表型。
所述特異性片段為300-500bp。
所述目的基因為綠盲蝽β-Actin,其序列如SEQ ID NO1所示。
所述目的基因β-Actin中合成dsRNA選定的特異性片段序列如SED ID NO2所示。
所述目的基因為綠盲蝽V-ATPaseE,其序列如SEQ ID NO3所示。
所述目的基因片段V-ATPaseE中合成dsRNA的選定特異性片段序列如SED ID NO4所示。
所述步驟(8)種,同時將GFP基因的dsRNA作為對照組混合到綠盲蝽人工飼料中,所述GFP基因序列如SEQ ID NO5所示。
所述GFP基因中選定合成dsRNA的靶標片段序列如SEQ ID NO6所示。
所述持續飼養時間為5-7天。
本發明用飼喂法對綠盲蝽進行RNAi實驗的飼喂參數(包括飼喂體積,飼喂dsRNA總量)進行確定,并挑選了綠盲蝽持家基因V-ATPaseE驗證綠盲蝽飼喂方法導入dsRNA能否引發RNAi效果并產生致死表型,結果表明導入V-ATPaseE基因dsRNA可以導致綠盲蝽生長發育受到嚴重影響,并進入體內后導致靶標害蟲快速死亡。
本發明首次建立了利用飼喂方法用于綠盲蝽基因功能研究的RNAi方案;同時利用建立的RNAi平臺篩選得到了持家基因、能量代謝相關基因、細胞凋亡相關基因等多個與綠盲蝽生長發育密切相關的基因。本研究為篩選新的抗蟲基因提供了新的技術手段和平臺,為發展抗綠盲蝽轉基因作物提供了新的基因資源。實踐表明,發展轉基因抗蟲作物能有效地控制害蟲危害,RNAi是由雙鏈RNA引發的基因沉默現象,其作用機制是通過阻止同源基因的翻譯或者轉錄而實現基因沉默。
附圖說明
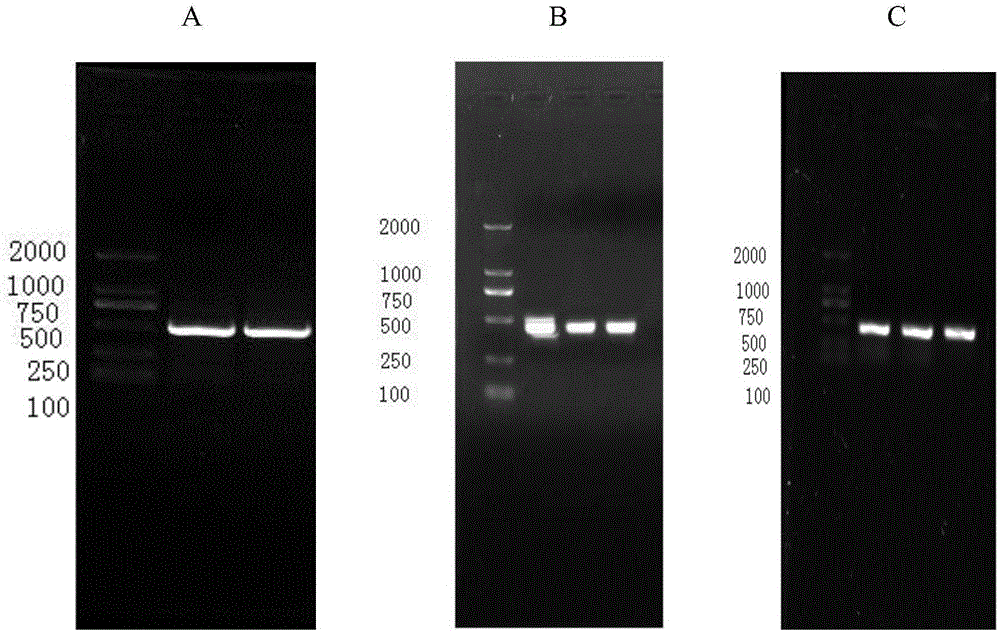
圖1實驗所用dsRNA凝膠電泳結果圖,
其中A、B、C分別表示為dsRNAβ-Actin、dsRNAGFP、dsRNAVA-E凝膠電泳結果。
圖2綠盲蝽飼喂方法現場圖片。
圖3β-Actin飼喂RNAi存活率。
圖4β-Actin飼喂RNAi后異常表型。
圖5飼喂dsV-ATPaseE濃度梯度對綠盲蝽存活率的影響。
圖6飼喂傳遞dsV-ATPaseEF1對綠盲蝽存活率的影響。
圖7飼喂傳遞dsV-ATPaseE F1對靶標基因表達量的影響。
具體實施方式
下面結合實施例對本發明作進一步的詳細說明。下面涉及到的化學試劑均為市售。
綠盲蝽的轉綠組測序和ApoV-ATPaseE基因的生物信息學分析。
本實驗室事前對綠盲蝽進行轉綠組測序,成功得到綠盲蝽63029條基因序列,并注釋了其中25760條基因序列。利用生物信息學技術及相關在線軟件對綠盲蝽轉錄組數據進行初步的分析,通過在線程序NCBI Blast等程序的幫助下確定了綠盲蝽ApoV-ATPaseE基因。
測序結果見SEQ ID NO:3所示。
1、綠盲蝽總RNA的提取
綠盲蝽總RNA提取的過程在超凈工作臺內進行(離心除外),以避免RNA酶(RNase)污染降解RNA。整個提取步驟如下所示:
1.取5頭經過72小時饑餓處理的綠盲蝽,加入經液氮冷卻的玻璃勻漿器中,立即向勻漿器內加入1ml RNA提取試劑(Trizol),將組織磨碎;將研磨好的組織溶液倒入1.5ml無RNA酶的離心管中,置于冰上;
2. 4℃,12000g離心15min,將上清液體轉移到新的1.5ml無RNA酶離心管中;
3.將上清在室溫下放置5min后,向離心管中加入200μl氯仿,用手劇烈震蕩15s,再室溫靜置2-3min;
4. 4℃,12000g離心15min,混合物在此時分層,下層為有機相,上層為水相(RNA在水相中);將上層水相轉移到新的1.5ml無RNA酶離心管中,向離心管中加入500μl異丙醇,室溫靜置10min;
5. 4℃,12000g離心10min后,棄掉上清(盡量吸凈),向離心管中加入1ml 75%的乙醇(用無RNA酶的H2O配置,現用現配),震蕩離心管以洗滌沉淀;
6. 4℃,7500g離心10min后,棄掉上清(盡量吸凈),在超凈工作臺中干燥5-10min;
7.加入20μl無RNA酶的H2O,震蕩離心,使沉淀充分溶解;
8. 60℃溫育10min后,取1μl樣品稀釋至5μl,其中2.5μl進行電泳檢測,2.5μl用濃度測量儀器(NanoDrop)檢測濃度;如果凝膠電泳檢測結果合格且樣品OD值如下:
260/280:1.80~2.00
260/230:1.80~2.00
則說明RNA質量良好,存于-80℃備用。
2、第1鏈cDNA的合成
整個反轉錄過程盡量保證在超凈工作臺內進行,操作步驟按第一鏈cDNA合成試劑盒進行,具體操作如下:
1.在新的1.5ml無RNA酶離心管中加入2μg RNA(根據濃度測量的結果計算需加入RNA的體積數),再加入1μl引物(oligo-dT),用無RNA酶的H2O補至12μl;65℃溫育5min,之后馬上置于冰上;
2.在離心管中分別加入下列試劑,使體系達到20μl:4μl的5×反應緩沖液;2μl的dNTP混合物(10mM);1μl的反轉錄酶和1μl的RNA酶抑制劑;簡單混勻后,離心片刻;
3. 42℃溫育1h,然后再70℃溫育5min;合成后存于-20℃冰箱,使用前稀釋10倍。
3、靶標基因的克隆
本發明選取了持家基因、能量代謝相關基因和細胞凋亡相關基因上的基因片段,GFP基因上的片段作為對照。持家基因又稱管家基因,是指所有細胞中均要表達的一類基因,其產物是對維持細胞基本生命活動所必需的;而能量代謝相關基因和細胞凋亡相關基因也在綠盲蝽生長發育和生理過程中起著非常重要的作用。本發明選取此類基因上的片段,利用飼喂法進行RNAi將其沉默,研究觀察沉默該基因后,對綠盲蝽個體發育造成的影響。GFP即綠色熒光蛋白,在昆蟲體內并不存在,故大量研究中將其作為對照基因合成dsRNA。
1.靶標基因引物的設計:
(1)利用BLAST檢索數據庫,搜索綠盲蝽的相近物種同一靶標基因的蛋白或者cDNA編碼的氨基酸序列,用綠盲蝽成蟲若蟲轉錄組組建的本地BLAST庫進行比對查詢,確定所選基因的序列。
(2)根據靶標基因的片段,用Primer Premier 5.0軟件設計帶T7啟動子的引物,引物設計原則如下:
a.引物設計的區間要在靶標基因的編碼區,中間片段大小在400~500bp左右。
b.中間片段盡量選在靶標基因的特異區域。
c.引物之間不能形成引物二聚體,交叉二聚體。
2.根據以上引物設計原則設計引物后,送至華大基因進行引物合成。
3.相關靶標基因的引物序列見表1-1。
表1.實驗所用dsRNA引物
4、目的片段的PCR擴增、測序及序列分析
1.以綠盲蝽cDNA為模板,用ExTaq酶擴增表1-1中基因。反應體系為25μl:17μl重蒸水,2.5μl 10×反應緩沖液,2μl dNTPs(2.5mM),1μl cDNA,1μl F引物(10mM),1μl R引物(10mM)和0.5μl DNA聚合酶。反應條件為:94℃預變性3min;94℃變性30s,60-65℃退火30s,72℃延伸30s,35個循環;最后72℃延伸10min。
2.PCR產物用溶解在1×TAE緩沖液的1%瓊脂糖凝膠電泳進行檢測。檢測正確的PCR產物,用瓊脂糖凝膠回收試劑盒進行回收(操作步驟按試劑盒內附帶的說明書進行),回收的片段連接到pGME-T載體上(體系為8μl膠回收產物、1μl 10×T4鏈接反應緩沖液、0.5μlpGME-T載體和0.5μl T4連接酶,16℃條件下連接過夜),連接體系轉化Top10感受態細胞(轉化步驟按感受態細胞附帶的說明書進行),培養過夜后,挑取8個陽性克隆進行PCR驗證(體系和條件同上),將檢測正確的克隆用液體LB培養基(含氨芐抗生素)培養過夜檢驗目的片段序列。
3.根據測序結果,將正確的菌株加入含有氨芐抗生素的新鮮LB培養基重新搖菌,獲得新鮮的菌液,將1ml菌液和百分之八十的甘油按照9:1比例混合,存于-80℃低溫冰箱保存。將剩余的菌液進行質粒提取。序列結果見序列表。
5、dsRNA模板的制備
以帶有靶標基因片段的質粒為模板,用聚合酶擴增。反應體系為25μl:17μl重蒸水,2.5μl10×反應緩沖液,2μl dNTPs(2.5mM),1μl載有靶標基因片段的質粒,1μl正向(F)引物(10mM),1μl反向(R)引物(10mM)和0.5μl DNA聚合酶。反應條件為:94℃預變性3min;94℃變性30s,55-60℃退火45s,72℃延伸1min,35個循環;最后72℃延伸10min。
將1μl PCR產物用溶解在1×TAE緩沖液的1%瓊脂糖凝膠電泳進行檢測。如果所得條帶長度與目的片段長度一致,并且結果顯示為單一明亮的條帶,則將剩余的PCR產物進行酚氯仿抽提與純化。
PCR產物的酚氯仿抽提的步驟:
1.將需要純化的PCR產物用無RNA酶的H2O定容至200μl,
2.加入等體積(200μl)的酚氯仿試劑(一定取酚氯仿試劑的下層,并且拿試劑瓶時要平穩,不能晃動)。
3.溫和混勻,在4℃離心機下離心(12000rpm,4℃)15min.
4.取上清,加入1/10體積(20μl)的3M乙酸鈉(pH5.2)和2倍體積(400μl)的100%乙醇(-20℃貯存),溫和混勻后放于-20℃靜置沉淀至少3小時或者過夜。
5.在4℃離心機下離心(12000rpm,4℃)30min.
6.白色沉淀積于離心管底部,棄上清液(為防止沉淀一起被吸出,可保留些許上清液),加入75%乙醇(-20℃貯存)輕輕混勻,洗滌沉淀
7.在4℃離心機下離心(7500rpm,4℃)5min.
8.將乙醇慢慢吸出(注意不要吸出沉淀),接近沉淀的殘留液體用10μl移液槍慢慢吸出。將離心管開蓋放入37℃恒溫培養箱干燥,10min左右,待乙醇全部揮發。
9.加入5μl RNase–free H2O輕彈管底,讓沉淀充分均勻溶解于無RNA酶的H2O。
10.取1μl溶液溶于4μl無RNA酶的H2O中,用濃度測量儀器檢測濃度和OD值。用凝膠電泳檢測產物單一性,如果檢測條帶是單一明亮的條帶,并且其OD值為:
260/280:1.8-2.0
260/230:1.8-2.0
則說明PCR產物質量良好,可以作為下一步合成dsRNA的模板使用。
6、dsRNA的合成
1.將ATP,CTP,GTP,UTP(100mM)放于冰上慢慢融化,5×反轉錄緩沖液室溫融化,T7RNA聚合酶放于-20℃貯存,隨用隨拿,用完立即放入-20℃環境下貯存。溶解以后輕彈管底,瞬時離心,將試劑離心到管底。
2.按以下比例將試劑混合:
輕彈混勻,瞬時離心。放入37℃金屬浴中4小時。
3.從金屬浴中取出離心管,放入75℃金屬浴中5min,然后放置室溫冷卻(切勿放在冰上)。吸出1μl,稀釋5倍,凝膠電泳檢測產物條帶。
4.去除DNA和dsRNA.按照比例加入下列試劑:
充分混勻,輕彈管底,瞬時離心后,放入金屬浴37℃30min,加入EDTA試劑1μl,65℃金屬浴中放置5min終止反應。取1μl產物,稀釋5倍,凝膠電泳檢測產物單一性,濃度檢測儀器檢測濃度和OD值。如果檢測條帶是單一明亮的條帶,并且其OD值為:
260/280:1.8-2.0
260/230:1.8-2.0
則說明dsRNA質量良好,可以進行下一步dsRNA酚氯仿抽提。
7、dsRNA的酚氯仿抽提
1.將需要純化的dsRNA(~60μl)用無RNA酶的H2O定容至200μl(可根據實際需要等比例擴大體系)。
2.加入1/2體積(100μl)的水飽和酚試劑(一定取水飽和酚試劑的下層,并且拿試劑瓶時要平穩,不能晃動)和1/2體積的氯仿(100μl)。
3.輕輕混勻,在4℃離心機下離心(12000rpm,4℃)15min。
4.取上層水相,加入的等體積氯仿(200μl),輕輕混勻,在4℃離心機下離心(12000rpm,4℃)15min。
5.取上層水相,加入1/10體積(20μl)的3M乙酸鈉(pH5.2)和2.5倍體積(500μl)的100%乙醇(-20℃貯存),溫和混勻后放于-20℃靜置沉淀至少3小時或者過夜。
6.在4℃離心機下離心(12000rpm,4℃)30min。
7.白色沉淀積于離心管底部,棄上清液(為防止沉淀一起被吸出,可保留些許上清液),加入80%乙醇(-20℃貯存)輕輕混勻,洗滌沉淀。
8.在4℃離心機下離心(7500rpm,4℃)5min。
9.將乙醇慢慢吸出(注意不要吸出沉淀),接近沉淀的殘留液體用10μl移液槍慢慢吸出。將離心管開蓋放入37℃恒溫培養箱干燥,10min左右,待乙醇全部揮發。
10.加入5μl無RNA酶的H2O輕彈管底,讓沉淀充分均勻的溶解于無RNA酶的H2O中。
11.取1μl溶解的dsRNA產物稀釋于4μl DEPC H2O中,用凝膠電泳檢測產物單一性,并用濃度檢測儀器檢測濃度和OD值。如果檢測的條帶是單一明亮的條帶,并且其OD值為:
260/280:1.8-2.0
260/230:1.8-2.0
則證明dsRNA質量良好,可以進行綠盲蝽體內注射RNAi實驗。
8、事實熒光定量PCR與分析
實驗采用的是2-ΔΔCT法進行相對定量,即通過內參基因校準不同樣品的模板加入總量的差異。ΔΔCT=(CT目的基因-CT內參基因)實驗組-(CT目的基因-CT內參基因)對照組,2-ΔΔCT的意義即為:實驗組的目的基因相對于對照組的目的基因的倍數,由此即可得到相對表達量。本實驗所用的引物如表2所示
表2實時熒光PCR實驗所用引物
本實驗中,以α-tubulin,GADPH為內參基因,按照TakaRa公司的SYBR PRemix Ex TaqTM試劑盒說明書對綠盲蝽對照組和實驗組進行Real Time PCR反應,以實驗的三組基因的第1d、2d、3d、4d、5d樣品的cDNA模板。反應體系以及反應條件如下:
反應體系為20μL
反應條件:
第1階段 預變性
95℃ 2min
第2階段 PCR擴增
95℃ 10s
55℃ 10s
第3階段:溶解曲線分析
溫度以0.5℃/s的速度從55℃增加到95℃,連續測定樣品的熒光強度以獲得溶解曲線。
9、3齡(3L)綠盲蝽的準備
本發明中所選綠盲蝽為3L若蟲,現以3L若蟲的收集方法如下:收集1L初孵若蟲(200~300頭),放入養蟲盒中,盒中放入干凈濾紙和一根去皮的新鮮玉米,約4天后所有1L若蟲成長為3L若蟲,準備鋪好濾紙和7~8粒新鮮玉米粒的塑料培養皿,每個培養皿里放15只3L若蟲,放若蟲時,用軟毛刷輕輕挑取,以免對若蟲造成不必要的機械損傷。饑餓處理兩個小時
人工飼料的配制
人工飼料是按照中國農業科學院植物保護研究所棉花害蟲研究組所提供的配方來配制的(棉蟲組已經在人工飼料的配制和實際應用方面通過了實驗驗證)。
利用β-Actin基因dsRNA綠盲蝽飼喂RNAi的存在驗證
人工飼料問題解決以后我們用管家基因dsRNA來驗證了綠盲蝽飼喂RNAi的存在與否,我們研究表明dsβ-Actin不但通過飼喂方法誘導RNAi而且有明顯的異常表型。結果如圖4所示。
10、綠盲蝽RNAi飼喂方法的具體實施
(1)實驗前準備;將以上所述的3L綠盲蝽,飼喂前每15個分一個培養皿,兩個小時饑餓處理。饑餓處理過程中把配好的人工飼料分到幾個離心管提前準備。把200ul的槍頭頭端用剪刀剪切1/3的部分,以便容易吸取粘稠的人工飼料。(2)將含有dsRNA的人工飼料用封口膜包起來(總體積200ul)輕混勻后喂給綠盲蝽。
飼喂蟲量為30-60頭為一組重復,至少重復三次,統計總數至少為120頭/基因。包裝完含有dsRNA的人工飼料后將裝有3L若蟲的培養皿放在光照培養箱內飼養,L:D=16:8,空氣濕度:55-60%,每天更換人工飼料,注意保持培養皿內清潔。每隔24小時統計每個培養皿中死亡蟲體數量,并做詳細記錄與統計。
1.最佳飼料傳遞方法和每次飼料體積的確定
在人工飼料的傳遞方法方面以前在刺吸式口器昆蟲和其他類昆蟲都有相關嘗試并取得了實際應用。本專利中我們為了降低dsRNA降解速度和實驗簡易,應用性等方面考慮,我們就應用了封口膜包飼料法(圖1),此方法方便簡易,操作簡單,所需時間不多等特點。
在本發明中另一個實驗前解決問題是確定每次飼喂量和飼喂數量(個體數量),考慮到重復試驗dsRNA的使用量和蟲子數量的最佳配合及高效和儉約性,我們首先使用了100ul,150ul和200ul飼料體積進行試驗,結果表明100ul和150ul雖然節約dsRNA量但試驗蟲子數量受限制。所以200ul飼料(培養皿)體積確定為下一步試驗飼料體積。
飼料體積確定以后下一步確定每培養皿/個體數量的確定。我們分別10頭,15頭和20頭三組做實驗。考慮到10頭/培養皿增加試驗量,20頭/培養皿卻因飼料量24h不夠而導致綠盲蝽饑餓死亡現象,所以我們把15頭/培養皿確定為下一步試驗個體數。
2.最佳dsRNA飼喂濃度的確定
在確定飼喂體積和個體數以后,本發明使用6種劑量的dsRNA V-ATPaseE:表1(0.01ul,0.03ul,0.1ul,0.3ul,0.5ul,0.8ul)的人工飼料和dsRNA的混合飼喂給3齡綠盲蝽若蟲飼喂后連續7天統計生存率,結果見表3所示,通過死亡率統計可以看出:隨著dsRNA濃度的提高綠盲蝽存活率明顯下降。0.3ul,0.5ul和0.8ul的濃度顯著提高了靶標害蟲的死亡率(P<0.05.LSD)。見圖5.
表3不同飼喂dsRNA濃度對綠盲蝽存活率的影響
考慮到今后飼喂實驗結果與dsRNA濃度的關系我們最后確定0.5ug/ul定位今后實驗的篩選濃度。
實施例1:飼喂篩選V-ATPaseE基因片段的dsRNA對綠盲蝽生長發育的影響
采用上述飼喂法用dsGFP和ds V-ATPaseE分別對3L綠盲蝽進行飼喂實驗,飼喂參數如下:飼喂體積為200ul,飼喂dsRNA最終濃度為0.5μg/μl。結果見圖6,7。
由圖6和圖7可以看出,處理組(dsV-ATPaseE)與對照組(dsGFP)存活率有明顯的區別,并且隨著天數的增加,兩組數值的差距逐漸變大。飼喂實驗后的第7天,對照組綠盲蝽仍有很高的存活率。據統計結果顯示,兩者具有顯著性差異(P<0.05)。
同理,采用本申請相同的方法可以將綠盲蝽體內與能量代謝相關基因、細胞凋亡相關基因進行沉默。由此看見,利用飼喂法對綠盲蝽進行RNA干擾(RNAi)能夠應用于基因篩選。
- 還沒有人留言評論。精彩留言會獲得點贊!