利用聚球藻UTEX2973生產糖類化合物的構建體、菌株與方法與流程
本發明涉及生物制造領域。具體而言是一種利用聚球藻utex2973生產糖類化合物的構建體、菌株與方法。
背景技術:
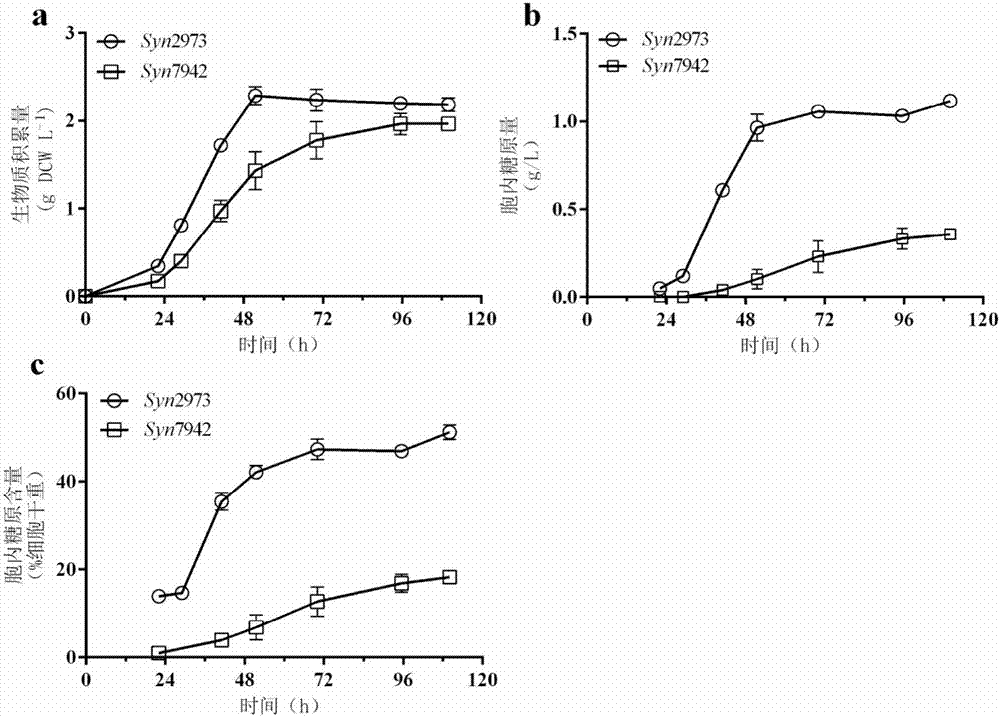
:代謝工程和合成生物學領域的快速發展,使得經過代謝工程改造的異養微生物,如大腸桿菌或酵母,已經可以將糖類原料發酵轉化成為許多種生物燃料(peralta-yahyaetal.2012)和生物基化學品(haraetal.2014)。與現有基于石化原料的化學品生產方式相比,基于糖原料的微生物發酵生產方式更環保且可持續,具有顯著的應用潛力。雖然目前微生物發酵生產化學品得到了廣泛的關注和發展,但是大部分產品的生產成本較高,目前僅有大約30種生物基化學品實現了商業化生產(e4techetal.2015)。而生物化學品生產的經濟可行性很大程度上依賴于糖原料成本。糧食作物如甘蔗、玉米等以及纖維素生物質都已經被成功地用作微生物發酵的原料(ziolkowska2014)。但是以糧食作物為原料用于微生物發酵存在了“與人爭糧、與糧爭地”的弊端,而且原料供應受限。纖維素生物質是自然界中含量最多的糖類物質,因此纖維素近年來被認為是具有前景的糖類原料(graham-rowe2011)。然而由于植物細胞壁中木質素和纖維素的降解和轉化成本較高,由纖維素作為原料的發酵產品與石油化工產品相比并沒有成本優勢(ziolkowska2014)。因此,廉價且可持續獲得的糖類原料對當今微生物發酵產業仍然十分重要,且對微生物發酵生產廉價化學品更為重要。藍細菌(藍藻)是一種能進行植物型放氧光合作用的原核微生物,是海洋生態系統的重要初級生產者(hernandez-prietoetal.2014)。與高等植物相比,藍細菌具有生長速度快、光合效率高、容易進行遺傳改造等優勢(zhouandli2010)。另外,水生的藍細菌可以用放置在不適宜耕種土地甚至漂浮在海洋中的培養器中進行培養。基于這些優勢,藍細菌已經在生產生物燃料、生物基化學品及糖類物質方面展現了其巨大的潛力,具有廣泛的應用前景(angermayretal.2015;guptaetal.2013;haysandducat2015;sarkarandshimizu2015)。蔗糖是微生物發酵生產生物乙醇的主要糖類原料,目前主要在甘蔗中提取(balatandbalat2009)。已知至少有60種藍細菌菌株可以合成蔗糖,蔗糖主要是作為藍細菌應對外界滲透脅迫的相容性物質(balatandbalat2009),也可作為穿梭于細胞之間的營養糖分以及信號分子(hagemann2011)。目前文獻報道的藍細菌生產蔗糖的研究工作主要以集胞藻pcc6803和聚球藻pcc7942作為宿主菌,通過代謝工程改造來提高其蔗糖產量(duetal.2013;ducatetal.2012)。通過在聚球藻pcc7942表達大腸桿菌來源的質子/蔗糖協同轉運蛋白cscb,研究人員得到了目前藍細菌生產蔗糖的最高產量36.1mg/l/h(ducatetal.2012)。根據研究人員估算,如果能夠將此工程菌大規模培養,其蔗糖年產量將是甘蔗的7倍(ducatetal.2012)。這些研究充分說明利用藍細菌細胞工廠生產蔗糖的巨大潛力。藍細菌中的糖原已經被證明可以作為糖類原料直接被微生物利用來生產乙醇(aikawaetal.2013)。除了作為藍細菌中主要的碳水化合物儲存物質(allen1984)和黑暗條件下呼吸作用的能量來源(grundeletal.2012),糖原還對藍細菌在缺氮、鹽脅迫及氧化脅迫的適應中起到重要作用(grundeletal.2012;hickmanetal.2013;suzukietal.2010)。aikawa等報道了他們利用聚球藻pcc7002生產糖原的工作,在氮源減少、補加氯化鈉的培養條件培養7天,聚球藻pcc7002胞內糖原積累速率為0.5g/l/d,這是目前文獻報道的藍細菌糖原最高產率(aikawaetal.2014)。由于糖原積累依賴于限制氮源的培養條件,大多數藍細菌糖原積累與細胞生物質積累之間存在明顯的矛盾(aikawaetal.2014)。因此,為了實現使用藍細菌生產糖原,需要一個能夠同時快速積累糖原和生物質的藍細菌菌株。聚球藻(synechococcuselongatus)utex2973,來源于美國德克薩斯大學藻種保藏中心(theculturecollectionofalgaeattheuniversityoftexasataustin),是目前已報道的生長速度最快的藍細菌(yuetal.2015)。雖然它與聚球藻pcc7942的基因組具有99.8%的同源性,在同樣的培養條件下聚球藻utex2973的生長速度是聚球藻pcc7942的兩倍,并且可以更好地耐受室外的高光、高溫環境(yuetal.2015)。在38℃、500μe/m2/s,通入3%co2的條件下,聚球藻utex2973的代時為2.3h,與酵母的生長代時相近(yuetal.2015)。技術實現要素:本發明的目的是一種利用聚球藻生產糖類化合物的構建體、菌株與方法。為實現上述目的,本發明采用技術方案為:一種利用聚球藻utex2973生產糖類化合物的構建體,構建體包含有在聚球藻utex2973中具有活性的啟動子,處于該啟動子控制之下的質子/蔗糖的協同轉運蛋白基因,用于篩選轉化體的抗生素抗性基因,以及用于基因組同源整合的同源臂。所述構建體含有所述在藍細菌中具有活性的啟動子上游的用于篩選藍細菌轉化體的抗生素抗性基因。所述質子/蔗糖的協同轉運蛋白基因為seqidno:1中所示的來源于大腸桿菌的cscb基因。所述抗生素抗性基因為氯霉素抗性基因,其例如顯示在seqidno:4中。所述在藍細菌中具有活性的啟動子選自plac啟動子。其它啟動子舉例:ppete,pcpcb,pcpt,penyc4等。其它抗生素抗性基因列舉:卡那霉素抗性基因,壯觀霉素抗性基因,慶大霉素抗性基因等適用于藍細菌的抗生素抗性基因。所述用于基因組同源整合的同源臂為來源于聚球藻utex2973的ns3基因的c-末端序列(seqidno:2)和n-末端序列(seqidno:3)。其他同源臂列舉:來源于聚球藻utex2973的ns1基因的c-末端序列(seqidno:12)和n-末端序列(seqidno:13);ns2基因的c-末端序列(seqidno:14)和n-末端序列(seqidno:15)。一種載體,其包含上述任一項的構建體。所述載體選自下列質粒,質粒psk015,其于2016年3月11日保藏于中國微生物菌種保藏管理委員會普通微生物中心,地址:北京市朝陽區北辰西路1號院3號,sk028,分類命名大腸埃希氏菌,escherichiacoli保藏號為cgmccno.12192。一種包含上述任一項的構建體的藍細菌。一種包含上述的載體轉化的藍細菌。所述藍細菌選自:于2016年3月11日保藏于中國微生物菌種保藏管理委員會普通微生物中心,地址:北京市朝陽區北辰西路1號院3號,藍細菌syn2973::cscb,分類命名為聚球藻synechococcussp.,其保藏號為cgmccno.12231。一種在藍細菌中生產糖類化合物的方法,在含有nacl或kcl的條件下培養任一項的藍細菌,從獲得的培養物中提取所需的糖類化合物。所述糖類化合物為蔗糖和糖原。所述藍細菌的培養條件為38℃,250μe/m2/s,通入3%co2,使用100-200mm的nacl或kcl對藍細菌進行鹽脅迫,于培養物細胞內外獲得所需的糖類化合物;優選為150mm。使用含有150mmkcl的培養基重懸藍細菌從而進行半連續培養,在培養21天之后共積累了8.1g/l的胞外蔗糖。本發明所具有的優點:本發明提供在光合微生物藍細菌體內利用太陽能固定二氧化碳合成糖類化合物,合成糖類物質的能量來自于太陽能,碳源來自于二氧化碳。因此,利用本發明方式制備的生物基產品不會受到原料不足的制約,使用這種生物基產品不會增加碳排放,是真正的零排放生物基產品。具體是,本發明在光合生物藍細菌聚球藻utex2973中合成糖類化合物構建體,包含所述構建體的載體,包含所述構建體或用所述載體轉化的藍細菌,以及在藍細菌中生產糖類化合物的方法。進而使得在藍細菌聚球藻utex2973內大量分泌蔗糖的同時也在胞內積累大量的糖原,分泌到胞外的蔗糖以及在胞內的糖原均可以回收用于微生物發酵。附圖說明圖1為本發明實施例提供的聚球藻utex2973與聚球藻pcc7942生物質和糖原含量對比圖;其中,a為兩個菌株的生物質積累情況;b為兩菌株在此培養條件下細胞內糖原的量;c為糖原分別占各自細胞干重的比例。圖2為本發明實施例提供的質粒psk015的基本結構;其中,在聚球藻utex2973的m744_rs12430基因上下游片段(ns3-n和ns3-c)之間有氯霉素抗性基因(cmr)、蔗糖轉運蛋白基因cscb以及plac啟動子。圖3為本發明實施例提供的加入iptg對syn2973::cscb蔗糖積累的影響圖;其中,a為是否加入iptg下的生長曲線;b為藍細菌胞外蔗糖的積累情況;c為胞內蔗糖的積累情況;d為胞外蔗糖占蔗糖總產量的百分比。圖4為本發明實施例提供的分別使用150mm的nacl或kcl鹽脅迫下聚球藻utex2973野生細胞的生物質積累(a)及胞內蔗糖積累情況(b)圖。圖5為本發明實施例提供的基因工程菌株syn2973::cscb在使用150mmkcl進行鹽脅迫下,半連續培養第一個生長周期胞外蔗糖積累情況(a)以及生物質分布情況(b)圖。圖6為本發明實施例提供的基因工程菌株syn2973::cscb在使用150mmkcl進行鹽脅迫下,半連續培養21天(7個周期)的生物質情況(a)及胞外蔗糖積累情況(b)圖。序列表信息:seqidno:1:來源于大腸桿菌(escherichiacoli)的蔗糖轉運蛋白(cscb)基因序列。seqidno:2:來源于聚球藻utex2973的ns3基因的c-末端序列。seqidno:3:來源于集聚球藻utex2973的ns3基因的n-末端序列。seqidno:4:質粒psk015上的氯霉素抗性序列(ncbiid:nc_005923.1)。seqidno:5:引物ns3-f的序列。seqidno:6:引物ns3-r的序列。seqidno:7:啟動子plac的序列。seqidno:8:啟動子ppete的序列。seqidno:9:啟動子pcpcb的序列。seqidno:10:啟動子pcpt的序列。seqidno:11:啟動子penyc4的序列。seqidno:12:來源于聚球藻utex2973的ns1基因的c-末端序列。seqidno:13:來源于集聚球藻utex2973的ns1基因的n-末端序列。seqidno:14:來源于聚球藻utex2973的ns2基因的c-末端序列。seqidno:15:來源于集聚球藻utex2973的ns2基因的n-末端序列。具體實施方式以下的實施例可以結合附圖使本領域的普通技術人員更全面地理解本發明,但不以任何方式限制本發明。相關術語藍細菌(也稱為藍藻)是一類光合自養的原核微生物,其能夠利用太陽能,固定二氧化碳為碳水化合物。質子/蔗糖的協同轉運蛋白(cscb)是能夠利用跨膜的質子梯度特異性地運輸蔗糖進出細胞的通道蛋白。在本發明的實施方案中,載體(vector)是指能夠將dna片段(目的基因)轉移到受者細胞中的一種自我復制的dna分子。接合轉移(conjugation)是指細菌細胞間通過直接細胞接觸或性鞭毛進行的遺傳物質的傳遞過程,適用于對非天然感受態的宿主細胞基因工程改造過程中。同源臂(homologyarms)是在對藍細菌遺傳改造的過程中,用于將啟動子、處于該啟動子控制之下的基因以及抗生素抗性基因,通過同源重組(homologousrecombination)整合到宿主藍細菌基因組的兩個dna片段;分為n-末端和c-末端。本發明的實施方案是利用在藍細菌中具有活性的啟動子驅動質子/蔗糖的協同轉運蛋白在藍細菌中表達,利用藍細菌光合生物的特點,吸收光能固定二氧化碳,在鹽脅迫條件下合成并分泌蔗糖;同時利用聚球藻utex2973生長速度快,可以在積累生物質的同時大量積累胞內糖原的特點,合成分泌蔗糖的同時在胞內積累糖原。實施例1:用聚球藻utex2973生產糖原1.實驗步驟(1)將聚球藻utex2973和pcc7942野生型藻株分別在柱式光反應器培養:普通玻璃管,柱高580mm,直徑30mm,裝液量150ml(可裝約300ml)。初始接種濃度od730為0.05,培養基為bg11培養基(bg11培養基成分為1.5g/lnano3,40mg/lk2hpo4·3h2o,36mg/lcacl2·2h2o,6mg/l檸檬酸,6mg/l檸檬酸鐵銨,1mg/ledta二鈉鹽,20mg/lnaco3,2.9mg/lh3bo3,1.8mg/lmncl2·4h2o,0.22mg/lznso4·7h2o,0.39mg/lnamoo4·2h2o,0.079mg/lcuso4·5h2o和0.01mg/lcocl2·6h2o);在38℃、250μe/m2/s光照、通入3%的co2空氣進行培養。在培養過程中,取樣測試od730和糖原含量。而藍細菌生物質(drycellweight,dcw,細胞干重)含量按1mlod730=1.0的藻細胞約相當于0.34mg換算。(2)糖原的檢測:將上述獲得藍細菌藻液離心后,用無菌水洗滌3次,重懸于30%(w/v)的koh,95℃溫浴2h,然后加入冷的乙醇至終濃度為70-75%(v/v),置于冰上至少2h,用于沉淀糖原。然后4℃,10000g離心10分鐘,糖原沉淀用70%和98%的乙醇洗滌兩次,60℃,10min真空離心干燥,糖原溶于100mm醋酸鈉溶液(ph4.5),加入糖化酶(novozyme,denmark)進行糖化,60℃,2小時,將糖原轉變為葡萄糖,然后通過測定葡萄糖含量計算糖原含量。2、實驗結果如圖1所示,在相同的培養條件(38℃、250μe/m2/s光照、通入3%的co2空氣)下,與聚球藻pcc7942相比,聚球藻utex2973的生物質積累速度達到1.56gdcw/l/d,比pcc7942快約50%(圖1a)。積累生物質的同時聚球藻utex2973糖原也迅速積累;在第22到51.5h,聚球藻utex2973糖原含量從細胞干重的14.7%上升到42.15%,糖原積累速率達到0.75g/l/d,高于目前文獻報道最高產率0.5g/l/d;最后在平臺期時其糖原含量達1.1g/l左右,占干重的51%以上。這些結果顯示了聚球藻utex2973更強的生物質和糖原積累能力,證明了其作為生物發酵糖原料的潛力。實施例2:構建用于導入聚球藻utex2973的載體psk015以ns3-f(5’-tcagccagctcgtcgtgatgtcgaaacccaagcc-3’)和ns3-r(5’-acagtcggcgtcacggcaacttgctccgcgtagg-3’)為引物對,以質粒pns3-3bwithcscb(ducatetal.2012)為模板進行pcr擴增;并將pcr產物,與經ecori/sali酶切,t4dna聚合酶補平的pbr322載體連接,從而得到重組質粒psk015(圖2)。pcr擴增的條件、體系:1、pcr擴增體系10×擴增緩沖液:5μl、4種dntp混合物(2.5mm):4μl、ns3-f(20mm):0.5μl、ns3-r(20mm):0.5μl、質粒模板dna(100ng/μl):0.5μl、fastpfuflydna聚合:1μl、加雙蒸水至50μl;2、pcr擴增條件:95℃5min、95℃30s、55℃30s、72℃3min(返回第二步,共30個循環)、72℃10min。實施例3:聚球藻utex2973的接合轉移以及接合子的篩選1、取1ml對數生長期(od730約為1.0)的聚球藻utex2973培養物,5000rpm離心5分鐘收集細胞;并用新鮮的bg11培養基洗滌細胞,再將細胞重懸于100μlbg11培養基中。2、在lb培養基(10g/l蛋白胨,10g/l氯化鈉,5g/l酵母抽提物)中分別培養含有prl443質粒和prl623質粒的大腸桿菌hb101菌株,以及含有psk015質粒的大腸桿菌hb101菌株的兩種菌株,將菌株培養至對數生長期(od600約為1.0);各取1ml大腸桿菌培養物,5000rpm離心5分鐘收集菌體;并用新鮮的lb培養基洗滌細胞,再將細胞分別重懸于100μllb培養基中。3、將含有prl443質粒和prl623質粒的大腸桿菌hb101菌株、含有psk015質粒的大腸桿菌hb101菌株、和聚球藻utex2973三種微生物細胞混合,均勻地涂布到鋪在bg11平板(未加抗生素)上的硝酸纖維素膜上,并置于30℃、30μe/m2/s光照條件下溫育12小時。4、將硝酸纖維素膜轉移到含有20μg/ml氯霉素的bg11平板上,并在30℃、30μe/m2/s的條件下繼續培養。5、大約4~5天,將接合子從平板上挑出,在新鮮的bg11平板(含有20μg/ml氯霉素)劃線;待細胞富集后,再將它們接入到液體bg11(含20μg/ml氯霉素)培養基中培養。6、將psk015質粒按照常規方式接合轉移到野生型聚球藻utex2973后,得到的藻株命名為syn2973::cscb。所述質粒psk015,其于2016年3月11日保藏于中國微生物菌種保藏管理委員會普通微生物中心,保藏號為cgmcc12192。實施例4:基因工程聚球藻utex2973合成并分泌蔗糖1、實驗步驟(1)將上述獲得基因工程藻株syn2973::cscb在柱式光反應器中培養;柱式光反應器為普通玻璃管,柱高580mm,直徑30mm,裝液量150ml(可裝液約300ml)。初始接種濃度od730為0.05,培養基為bg11(補加20μg/ml氯霉素),在38℃、250μe/m2/s光照、通入3%的co2空氣的條件下進行培養。(2)藍細菌鹽脅迫:向上述步驟培養至處于對數末期的藍細菌培養基中添加使用bg11溶液配制的5mnacl,使得培養基中的nacl濃度為150mm。同時,向藍細菌培養物中加入iptg(異丙基硫代半乳糖苷)至終濃度為1mm;而以不加入iptg的上述培養物作為對照。鹽脅迫之后,連續取樣監測藍細菌細胞內外蔗糖含量。(3)藍細菌胞內和胞外蔗糖提取:取1ml上述鹽脅迫之后藍細菌細胞藻液,8000rpm離心10min收集藻細胞。離心所得上清液,經稀釋后用離子色譜檢測,即得到細胞外蔗糖含量。向細胞沉淀加入1ml80%乙醇混勻,65℃水浴4h。8000rpm離心10min,棄沉淀,上清用n2吹干,加入合適的ddh2o稀釋后,使用離子色譜進行濃度測定,即得到細胞內蔗糖含量。(4)離子色譜檢測蔗糖含量:使用離子色譜isc-3000(dionex,usa),分析柱:ma1(dionex,usa)。流動相為800mmnaoh溶液,流速0.4ml/min,蔗糖出峰約在28min處。2、實驗結果(1)由上述可見使用150mmnacl對syn2973::cscb藻株進行鹽脅迫,同時比較了iptg加入對蔗糖產量的影響。結果發現,在不加入iptg時,細胞的生長狀態更好(圖3a),并且胞外可以積累達3.2g/l的蔗糖(圖3b)。在使用1mmiptg誘導時,鹽脅迫下細胞生長狀態變差(圖3a),同時蔗糖產量降低為約2g/l(圖3b)。而比較兩種條件下藍細菌細胞內的蔗糖含量發現,加入iptg后,藍細菌胞內蔗糖含量相對于對照顯著降低(圖3c)。通過計算發現,即使不加入iptg,syn2973::cscb藻株產生的蔗糖也有超過94%被分泌到細胞外;加入iptg,藻株蔗糖分泌的比例與對照相比稍高(圖3d)。這一結果說明,導入cscb基因的藻株能夠非常有效地將其在鹽脅迫中積累的蔗糖分泌到胞外。實施例5:相同濃度氯化鈉和氯化鉀對聚球藻utex2973蔗糖合成的影響1、實驗步驟(1)將聚球藻utex2973野生型藻株在柱式光反應器中培養;柱式光反應器為普通玻璃管,柱高580mm,直徑30mm,裝液量150ml(可裝液約300ml)。初始接種濃度od730為0.05,培養基為bg11,在38℃、250μe/m2/s光照、通入3%的co2空氣的條件下進行培養。(2)藍細菌鹽脅迫:向上述培養至處于對數末期的野生型藍細菌培養基中添加使用bg11溶液配制的5mnacl或3mkcl,使得培養基中的鹽濃度為150mm。鹽脅迫之后,連續取樣監測藍細菌細胞內外蔗糖含量。(3)藍細菌胞內蔗糖提取和離子色譜檢測蔗糖含量的方法參照實施例4。2、實驗結果(1)由上述可見分別使用150mm的nacl和kcl對聚球藻utex2973野生型進行鹽脅迫,對比其胞內、胞外蔗糖的積累情況。實驗發現,聚球藻utex2973野生型藻株在150mmkcl脅迫下的生長狀態明顯好于150mmnacl脅迫(圖4a),并且兩種鹽脅迫下細胞生產蔗糖的能力并沒有明顯區別(圖4b)。這一實驗結果說明kcl也能夠誘導藍細菌合成積累蔗糖,而kcl脅迫時藍細菌生長狀態更好。實施例6:半連續培養模式下syn2973::cscb藻株的蔗糖合成1、實驗步驟(1)將上述實施例獲得的基因工程藻株syn2973::cscb在柱式光反應器中培養;柱式光反應器為普通玻璃管,柱高580mm,直徑30mm,裝液量150ml(可裝液約300ml)。初始接種濃度od730為0.05,培養基為bg11(補加20μg/ml氯霉素),在38℃、250μe/m2/s光照、通入3%的co2空氣的條件下進行培養。(2)藍細菌鹽脅迫:向上述步驟培養至處于對數末期的藍細菌培養基中添加使用bg11溶液配制的3mkcl,使得培養基中的kcl濃度為150mm,在鹽脅迫下繼續培養。(3)之后,每3天重復如下操作:8000rpm離心10分鐘收集藍細菌細胞;然后將細胞重懸于等體積的含有150mmkcl和20μg/ml氯霉素的新鮮bg11培養基中,繼續下一個培養周期(3天)。每個培養周期結束時,取樣用于監測細胞外蔗糖含量。(4)離子色譜檢測蔗糖含量的方法參照實施例4。2、實驗結果在150mmkcl脅迫條件下,syn2973::cscb胞外蔗糖含量以24.64mg/l/h的速率線性增加(圖5a)。在kcl脅迫后第54.5h,syn2973::cscb向細胞外分泌的蔗糖含量占細胞干重的81.43%;與此同時,胞內糖原含量占細胞干重的41.83%,這一比例與野生型菌株在nacl脅迫相同時間后的糖原含量相當。這一結果表明,即使syn2973::cscb在鹽脅迫條件下合成并分泌了大量的蔗糖,其胞內糖原含量仍然可以維持一定的比例;這顯示了聚球藻syn2973::cscb強大的糖類化合物合成能力。而該藻株的這一特點,使得將來利用該藻株生產蔗糖的同時,藍細菌細胞生物質由于富含糖原也可以用作微生物發酵的原料。在半連續培養模式下,從第2個培養周期開始syn2973::cscb細胞生長狀況得到明顯改善;在經過7個培養周期(21天),syn2973::cscb生長依舊良好(圖6a)。從蔗糖分泌的情況看,在前3個培養周期,蔗糖產量保持穩定,約為2g/l;雖然隨后單位培養周期蔗糖產量出現下降趨勢,但是在7個培養周期(21天)內總共分泌了約8.70g/l的蔗糖。這些結果進一步展示了聚球藻syn2973::cscb強大的糖類化合物合成能力。生物材料樣品保藏信息菌株保藏號保藏時間大腸桿菌sk028,其包含質粒psk015cgmcc121922016年3月11日藍細菌syn2973::cscbcgmcc122312016年3月11日上述菌株均保藏于中國微生物菌種保藏管理委員會普通微生物中心(chinageneralmicrobiologicalculturecollectioncenter,cgmcc)另外,上述實例中的啟動子抗生素抗性基因、同源臂分別可由下述記載的啟動子、抗生素抗性基因和同源臂替換;啟動子為ppete,pcpcb,pcpt,penyc4等。抗生素抗性基因為卡那霉素抗性基因,壯觀霉素抗性基因,慶大霉素抗性基因等適用于藍細菌的抗生素抗性基因。同源臂為來源于聚球藻utex2973的ns1基因的c-末端序列和n-末端序列、來源于聚球藻utex2973的ns2基因的c-末端序列和n-末端序列。參考文獻aikawas,josepha,yamadar,izumiy,yamagishit,matsudaf,kawaih,changjs,hasunumat,kondoa(2013)directconversionofspirulinatoethanolwithoutpretreatmentorenzymatichydrolysisprocesses.energenvironsci6(6):1844-1849.doi:10.1039/c3ee40305jaikawas,nishidaa,hosh,changjs,hasunumat,kondoa(2014)glycogenproductionforbiofuelsbytheeuryhalinecyanobacteriasynechococcussp.strainpcc7002fromanoceanicenvironment.biotechnolbiofuels7:88.doi:10.1186/1754-6834-7-88allenmm(1984)cyanobacterialcellinclusions.annurevmicrobiol38(1):1-25.doi:10.1146/annurev.mi.38.100184.000245angermayrsa,roviraag,hellingwerfkj(2015)metabolicengineeringofcyanobacteriaforthesynthesisofcommodityproducts.trendsbiotechnol33(6):352-361.doi:10.1016/j.tibtech.2015.03.009balatm,balath(2009)recenttrendsinglobalproductionandutilizationofbio-ethanolfuel.applenerg86(11):2273-2282.doi:10.1016/j.apenergy.2009.03.015duw,liangfy,duanyk,tanxm,luxf(2013)exploringthephotosyntheticproductioncapacityofsucrosebycyanobacteria.metabolicengineering19:17-25.doi:doi10.1016/j.ymben.2013.05.001ducatdc,avelar-rivasja,wayjc,silverpa(2012)reroutingcarbonfluxtoenhancephotosyntheticproductivity.appliedandenvironmentalmicrobiology78(8):2660-2668.doi:10.1128/aem.07901-11e4tech,re-cord,wur(2015)fromthesugarplatformtobiofuelsandbiochemicals.finalreportfortheeuropeancommission:contractno.ener/c2/423-2012/si2012.673791.graham-rowed(2011)agriculture:beyondfoodversusfuel.nature474(7352):s6-s8.doi:10.1038/474s06agrundelm,scheunemannr,lockauw,zilligesy(2012)impairedglycogensynthesiscausesmetabolicoverflowreactionsandaffectsstressresponsesinthecyanobacteriumsynechocystissp.pcc6803.microbiology158(12):3032-3043.doi:10.1099/mic.0.062950-0guptav,rathask,sooda,chaudharyv,prasannar(2013)newinsightsintothebiodiversityandapplicationsofcyanobacteria(blue-greenalgae)—prospectsandchallenges.algalres2(2):79-97.doi:10.1016/j.algal.2013.01.006hagemannm(2011)molecularbiologyofcyanobacterialsaltacclimation.femsmicrobiolrev35(1):87-123.doi:10.1111/j.1574-6976.2010.00234.xharaky,arakim,okain,wakais,hasunumat,kondoa(2014)developmentofbio-basedfinechemicalproductionthroughsyntheticbioengineering.microbialcellfactories13(1):1-19.doi:10.1186/s12934-014-0173-5hayssg,ducatdc(2015)engineeringcyanobacteriaasphotosyntheticfeedstockfactories.photosynthesisres123(3):285-295.doi:10.1007/s11120-014-9980-0hernandez-prietoma,semeniukta,futschikme(2014)towardasystems-levelunderstandingofgeneregulatory,proteininteraction,andmetabolicnetworksincyanobacteria.frontgenet5:191.doi:10.3389/fgene.2014.00191hickmanjw,kotovickm,millerc,warrenerp,kaiserb,juristat,buddem,crossf,robertsjm,carletonm(2013)glycogensynthesisisarequiredcomponentofthenitrogenstressresponseinsynechococcuselongatuspcc7942.algalres2(2):98-106.doi:10.1016/j.algal.2013.01.008peralta-yahyapp,zhangf,delcardayresb,keaslingjd(2012)microbialengineeringfortheproductionofadvancedbiofuels.nature488(7411):320-328.doi:10.1038/nature11478sarkard,shimizuk(2015)anoverviewonbiofuelandbiochemicalproductionbyphotosyntheticmicroorganismswithunderstandingofthemetabolismandbymetabolicengineeringtogetherwithefficientcultivationanddownstreamprocessing.bioresourcesandbioprocessing2(1):17.doi:10.1186/s40643-015-0045-9suzukie,ohkawah,moriyak,matsubarat,nagaikey,iwasakii,fujiwaras,tsuzukim,nakamuray(2010)carbohydratemetabolisminmutantsofthecyanobacteriumsynechococcuselongatuspcc7942defectiveinglycogensynthesis.applenvironmicrobiol76(10):3153-3159.doi:10.1128/aem.00397-08yuj,libertonm,cliftenpf,headrd,jacobsjm,smithrd,koppenaaldw,brandjj,pakrasihb(2015)synechococcuselongatusutex2973,afastgrowingcyanobacterialchassisforbiosynthesisusinglightandco2.scirep5:8132.doi:10.1038/srep08132zhouj,liy(2010)engineeringcyanobacteriaforfuelsandchemicalsproduction.proteincell1(3):207-210.doi:10.1007/s13238-010-0043-9ziolkowskajr(2014)prospectivetechnologies,feedstocksandmarketinnovationsforethanolandbiodieselproductionintheus.biotechnolrep4:94-98.doi:10.1016/j.btre.2014.09.001序列表seqidno:1大腸桿菌(escherichiacoli)的蔗糖轉運蛋白(cscb)基因序列atggcactgaatattccattcagaaatgcgtactatcgttttgcatccagttactcatttctcttttttatttcctggtcgctgtggtggtcgttatacgctatttggctgaaaggacatctagggttgacagggacggaattaggtacactttattcggtcaaccagtttaccagcattctatttatgatgttctacggcatcgttcaggataaactcggtctgaagaaaccgctcatctggtgtatgagtttcatcctggtcttgaccggaccgtttatgatttacgtttatgaaccgttactgcaaagcaatttttctgtaggtctaattctgggggcgctcttttttggcctggggtatctggcgggatgcggtttgcttgacagcttcaccgaaaaaatggcgcgaaattttcatttcgaatatggaacagcgcgcgcctggggatcttttggctatgctattggcgcgttctttgccggcatattttttagtatcagtccccatatcaacttctggttggtctcgctatttggcgctgtatttatgatgatcaacatgtgttttaaagataaggatcaccagtgcgtagcggcggatgcgggaggggtaaaaaaagaggattttatcgcagttttcaaggatcgaaacttctgggttttcgtcatatttattgttgggacgtggtctttctataacatttttgatcaacaactttttcctgtcttttatgcaggtttattcgaatcacacgatgtaggaacgcgcctgtatggttatctcaactcattccaggtggtactcgaagcgctgtgcatggcgattattccgttctttgtgaatcgggtagggccaaaaaatgcattacttatcggtgttgtgattatggcgttgcgtatcctttcctgcgcgctgttcgttaacccctggattatttcattagtgaagctgttacatgccattgaggttccactttgtgtcatatccgtcttcaaatacagcgtggcaaactttgataagcgcctgtcgtcgacgatctttctgattggttttcaaattgccagttcgcttgggattgtgctgctttcaacgccgactgggatactctttgaccacgcaggctaccagacagttttcttcgcaatttcgggtattgtctgcctgatgttgctatttggcattttcttcctgagtaaaaaacgcgagcaaatagttatggaaacgcctgtaccttcagcaatatag(a)序列特征:●長度:1248堿基對●類型:脫氧核糖核酸(dna)●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:大腸桿菌seqidno:2聚球藻utex2973的m744_rs12430基因的c-末端序列(也包括一部分基因上游序列)(ncbiid:nz_cp006471.1)tcagccagctcgtcgtgatgtcgaaacccaagccaccctcagaggtgaaggccgcttcgagcacatcgggaagcgtgtcgacgacggtgctcggtgcctcgggtaagagccagatagatgcggtgacgttcaccgtcgcgatcgtggcgggaaccacggtcactgcgtccgttacaactcgcacgtcgtcggccagcacgactgactcgacggcttcgattagcccgggagaggcaaggccatcaggctcggtagacagaatgctgattaacacctcgccgggagcagggctgctcacagccgcgtccttcacccgtgggtcagccgtcagcgcttgatagcggtaccaggccgcaccgcctgcggtcgagctgcccttgatccgctcgatggtgcgatcgcgaagctcgccatccggctcattcggctgccgcaccacgccgtagaacgcggcaaggttgtcgaggtcggggccgtttgagtagcgaagcagtgtcgcaagaagtgcgtcgttgatccgctgacgcaggatcagctcgcgggctgcgcagacctccagcagcttgatgaccgggtcggattcgaggatggcggtgtagctggcgtcacgcgatcgcaggtcgtcgatcaagtcctgcaggatcagttcaaagtccagcgcttcgatgatggtgggcgcgggaatcgtagcaaagtcaagaacggtcatgagacgactaagccctccagcgtgatacgcctgccctcggggatgtagtagccgatcaggttcagctcgacttgaccagctgcgctggctgagacgatgcgaaccttctccagcttcagccgtggctcccagcgatcgagcgcttcagctgtggccgccaccaggtcaacgatgagggactggttgatcgg(a)序列特征:●長度:900堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973seqidno:3:聚球藻utex2973的m744_rs12430基因的n-末端序列(也包括一部分基因下游序列)(ncbiid:nz_cp006471.1)gacaagccggggcagacgtgagccgtagtcccgtcgccagacgcgggtgcccacgggcgtcgtcaggatgtccgtaattgactgccggaggtggtcaatgcccttcagctccttgccactgtcacggctcatgcctcgggtcattagtcgcccgctccggtatcttcactggcttcgatgattgccgccccgcagctgcagaggtcaccgatccgagcagtcggcctctggttggtaaagaccgtgcgactgccggtgatgatcgtgttcaagccatgcagggggcaggcgtgaaggtcatcctttcgggccacggggcggctgttgacgaaggtgtcgtcgctcccggtgatgatgatcccgccgtgatcggtcacgtcgtttagtcgagcgatgcctggcgtcgtagtcacgggtttaggtcaatacgacttgcggtcactgtaacgttgccctcggcggtcacgttaacgtcgccttgggcttcgacttgcgcctcctgcacaaggatcacaatccgtccttgggctgcggtgaggtcgatcttgtactcatgcgcttcgcggtcgtactggatgattgagtcatcctcgaactgcgtcttttggatcgtttctttgtcctcgatctgggggtagtcagtcgagaacgcgccgggcatcgcgaagccctgactgatctcgccggagggggccatcacgacgacggcctcaccgacctcgggcgcccaccagaaccgatccttgcccgctcgctgcgtgagccacggaatccagtcagtgaggagcagcgcctcgccgctctcctcgtcttcctcgatcgcgacacggatcagccccttgggatagtcagcctcggctaccctgcctacgcggagcaagttgccgtgacgccgactgt(a)序列特征:●長度:900堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973seqidno:4:氯霉素抗性序列atggagaaaaaaatcactggatataccaccgttgatatatcccaatggcatcgtaaagaacattttgaggcatttcagtcagttgctcaatgtacctataaccagaccgttcagctggatattacggcctttttaaagaccgtaaagaaaaataagcacaagttttatccggcctttattcacattcttgcccgcctgatgaatgctcatccggagttccgtatggcaatgaaagacggtgagctggtgatatgggatagtgttcacccttgttacaccgttttccatgagcaaactgaaacgttttcatcgctctggagtgaataccacgacgatttccggcagtttctacacatatattcgcaagatgtggcgtgttacggtgaaaacctggcctatttccctaaagggtttattgagaatatgtttttcgtctcagccaatccctgggtgagtttcaccagttttgatttaaacgtggccaatatggacaacttcttcgcccccgttttcactatgggcaaatattatacgcaaggcgacaaggtgctgatgccgctggcgattcaggttcatcatgccgtctgtgatggcttccatgtcggcagaatgcttaatgaattacaacagtactgcgatgagtggcagggcggggcgtaa(a)序列特征:●長度:660堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:質粒pns3-3bwithcscbseqidno:5:引物ns3-f的序列tcagccagctcgtcgtgatgtcgaaacccaagccseqidno:6:引物ns3-r的序列acagtcggcgtcacggcaacttgctccgcgtaggseqidno:7:啟動子plac的序列atcaaataaaacgaaaggctcagtcggaagactgggcctttcgttttatctgttgtttgtcggtgaacgctctcctgagtaggacaaatccgccgggagcggatttgaacgttgcgaagcaacggcccggagggtggcgggcaggacgcccgccataaactgccaggcatcaaattaagcagaaggccatcctgacggatggcctttttgcgtttctactctagtttcgaaaccccaggcttgacactttatgcttccggctcgtataatgtgtggaattgtgagcggataacaatttcacacaaggaggaaaaacatatgtctaga(a)序列特征:●長度:327堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:pns3-3bwithcscbseqidno:8:啟動子ppete的序列aaggattcatagcggttgcccaatctaactcagggagcgacttcagcccacaaaaaacaccactgggcctactgggctattcccattatcatctacattgaagggatagcaagctaatttttatgacggcgatcgccaaaaacaaagaaaattcagcaattaccgtgggtagcaaaaaatccccatctaaagttcagtaaatatagctagaacaaccaagcattttcggcaaagtactattcagatagaacgagaaatgagcttgttctatccgcccggggctgaggctgtataatctacgacgggctgtcaaacattgtgataccatgggcagaagaaaggaaaaacgtccctgatcgcctttttgggcacggagtagggcgttaccccggcccgttcaaccacaagtccctatagatacaatcgccaagaagt(a)序列特征:●長度:435堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:集胞藻pcc6803seqidno:9:啟動子pcpcb的序列ttcgttataaaataaacttaacaaatctatacccacctgtagagaagagtccctgaatatcaaaatggtgggataaaaagctcaaaaaggaaagtaggctgtggttccctaggcaacagtcttccctaccccactggaaactaaaaaaacgagaaaagttcgcaccgaacatcaattgcataattttagccctaaaacataagctgaacgaaactggttgtcttcccttcccaatccaggacaatctgagaatcccctgcaacattacttaacaaaaaagcaggaataaaattaacaagatgtaacagacataagtcccatcaccgttgtataaagttaactgtgggattgcaaaagcattcaagcctaggcgctgagctgtttgagcatcccggtggcccttgtcgctgcctccgtgtttctccctggatttatttaggtaatatctctcataaatccccgggtagttaacgaaagttaatggagatcagtaacaataactctagggtcattactttggactccctcagtttatccgggggaattgtgtttaagaaaatcccaactcataaagtcaagtaggagattaat(a)序列特征:●長度:591堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:集胞藻pcc6803seqidno:10:啟動子pcpt的序列ttaacaaaaaagcaggaataaaattaacaagatgtaacagacataagtcccatcaccgttgtataaagttaactgtgggattgcaaaa(a)序列特征:●長度:88堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:集胞藻pcc6803seqidno:11:啟動子penyc4的序列aaatattctgaaatgagctgttgacaattaatcatccggctcgtataatgtgtggaattgtgagcggataacaatttcatacgctcacaattggtaccggtgataccagcatcgtcttgatgcccttggcagcaccctgctaaggaggcaacaag(a)序列特征:●長度:155堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:人工合成dnaseqidno:12:來源于聚球藻utex2973的ns1基因的c-末端序列。tccctgctcgtcacgctttcaggcaccgtgccagatatcgacgtggagtcgatcactgtgattggcgaaggggaaggcagcgctacccaaatcgctagcttgctggagaagctgaaacaaaccacgggcattgatctggcgaaatccctaccgggtcaatccgactcgcccgctgcgaagtcctaagagatagcgatgtgaccgcgatcgcttgtcaagaatcccagtgatcccgaaccataggaaggcaagctcaatgcttgcctcgtcttgaggactatctagatgtctgtggaacgcacatttattgccatcaagcccgatggcgttcagcggggtttggtcggtacgatcatcggccgctttgagcaaaaaggcttcaaactggtgggcctaaagcagctgaagcccagtcgcgagctggccgaacagcactatgctgtccaccgcgagcgccccttcttcaatggcctcgtcgagttcatcacctctgggccgatcgtggcgatcgtcttggaaggcgaaggcgttgtggcggctgctcgcaagttgatcggcgctaccaatccgctgacggcagaaccgggcaccatccgtggtgattttggtgtcaatattggccgcaacatcatccatggctcggatgcaatcgaaacagcacaacaggaaattgctctctggtttagcccagcagagctaagtgattggacccccacgattcaaccctggctgtacgaataaggtctgcattccttcagagagacattgccatgcccgtgctgcgatcgcccttccaagctgccttgccccgctgtttcgggctggcagccctggcgttggggctggcgaccgcttgccaagaaagcagcgctccgccggctgccggatcccagcccttgggcttggcgatcgctcaaaccggcaatgcctctctgtttggtcaggaacagttggcgggtcttcagattgccaaagcagtctttgaacgacctgctactggtgttccgattgactttcgcttgcaggacagtggcagtgatga(a)序列特征:●長度:1042堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973seqidno:13:來源于集聚球藻utex2973的ns1基因的n-末端序列。tggatgtgatcggaaccctgagccgtcgtcctggaggtgaagtctaatgtttgaaacgatttttgcgctgctgattgttctaggcgctggcgccggggctggcagcttagtcctgcgcaatctctactacatctgccaacccagtgaaattttgatctttgctggcagtagtcgccgcagtagtgatggccgccgagttggctatcgcttggtcaagggcggcagcagcctgcgggtacctctgctggaaaaagcgctccgcatggatctgaccaacatgatcattgagttgcgcgtttccaatgccttctccaagggcggcattcccctgactgttgaaggcgttgccaatatcaagattgctggggaagaaccgaccatccacaacgcgatcgagcggctgcttggcaaaaaccgtaaggaaatcgagcaaattgccaaggagaccctcgaaggcaacttgcgtggtgttttagccagcctcacgccggagcagatcaacgaggacaaaattgcctttgccaaaagtctgctggaagaggcggaggatgaccttgagcagctgggtctagtcctcgatacgctgcaagtccagaacatttccgatgaggtcggttatctctcggctagtggacgcaagcagcgggctgatctgcagcgagatgcccgaattgctgaagccgatgcccaggctgcctctgcgatccaaacggccgaaaatgacaagatcacggccctgcgtcggatcgatcgcgatgtagcgatcgcccaagccgaggccgagcgccggattcaggatgcgttgacgcggcgcgaagcggtggtggccgaagctgaagcggacattgctaccgaagtcgctcgtagccaagcagaactccctgtgcagcaggagcggatcaaacaggtgcagcagcaacttcaagccgatgtgatcgccccagctgaggcagcttgtaaacgggcgatcgcggaagcgcggggggccgccgcccgtatcgtcgaagatggaaaagctcaagcggaagggacccaacggctggcggaggcttggcagaccgctggtgctaatgcccgcgacatcttcctgctccagaag(a)序列特征:●長度:1099堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973seqidno:14:來源于聚球藻utex2973的ns2基因的c-末端序列。ctgaaacggtgaagtttgcattgtttttaagcacgagccatcaacagtaagagcgatcgcgctgggacgatgtaatcgcgccgtaggcagggttttgcctcatccggcagatgacaagcttcaagattcggcagtgaagtatcgagcaagcgataccaaacgcggccgcgaggaggcctcggcaactggaagcgcaggtcttcccagtaagcattaaaggctaggtaaagccattcctgctggcgaggatggcagagactgacggccagactgtgggaccacagcgcccaatcgggttgtttgagtttgacgccatgccagatggcatagggacgacgcggatggggttcgttctgcagcagttcgttctgttggaacatcaccagcgactgggaaagttcaatcaggcggcgactgaacaccaagaaatcggcatggcgatcgcacagcgaccaatcaaaccagctgatctcattgtcttggcagtaggcgttattgttaccctgctgactgcgtttgacctcatcgcccatcgtcagcatcggtgtgccctgggcgaggaataacgtggcgagcaaattgcgctgctgccgttcccgtaagctcagaatcgtggggtcatcggtctcgccttcaatgccgtagttccagctgtagttgtcattggtcccgtcccgattgttctctccattggcaaagttgtgcttctggctatagctgactagatctcgcagcgtaaagccgtcatggcaggtgatgaagttaatggtgcgtccggcataccattggtctgtgctgtagacatcggggctacccagcaggcgttgactgagggcgtaagtacagccctgatctccacgccaaaaacgccgaatatcgtcccggaagggaccgttccaagtcccaa(a)序列特征:●長度:907堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973seqidno:15:來源于集聚球藻utex2973的ns2基因的n-末端序列。acggcagcgaaagggcgacggcgcgatcgccggtatggaaaacgcttgctgcagcatcagatcgatgcgatcgccgaagactgtgacctagatgaggaagcatgagcgaactgggcttgagtctgacggcgatcgcgatttttaccacgacggcattagctttggtgggaccaatgctgggtagttctccgctgctaccggcgggattgggttttagcctcttggtgctgttcagtctggatgcggtgacttggcaggggcggggtgccacgttactgctcgatggcattcagcagcgatcgcccgaatatcgtcagcggattttgcatcacgaagcgggtcactacttggtagcaaccgcgctggggttacccgtgacgggctacaccctctcagcgtgggaagcgctgcgccaaggacaacctggtcgcgggggtgtgcagttccaagcagctgcgctagaagccgaagccgcacaggggcaactcagtcagcgatcgctggaacagtggtgtcaggtgttgatggccggtgcagcggcagagcaactggtctacggcaacgtggaagggggagctgacgatcgcgcccagtggaaacaactgtggcggcaactcgatcgcaatcctgccgaagcggatttacgcagtcgctggggattgttacgggcgaagactttactggagcaacaacgtcccgcctacgatgctttggtggcggcgatggctgcagaggccagcattgaagactgcaatcaagcgatcgccactgcttgggtagaagaacctgcgatcgccgcttagtgaagagtccagaagattcccctcccctctcgcccaatggacaagggaaatgtgattcagcatgagtcaagtccccagtgcatggatgggtgtgcaatgacggacagaggctgttgactctcagttatccctgtaagccaaatcgtccgaaaatcaccagctgaaacggttagggttgcattgtttttaagcacgagccatcaacagtaagagcgatcgcgcaggaacggtttagtgtgtagttgcgg(a)序列特征:●長度:1051堿基對●類型:dna●鏈型:單鏈●拓撲結構:線性(b)分子類型:dna(c)假設:否(d)反義:否(e)最初來源:聚球藻utex2973。當前第1頁12
相關技術
網友詢問留言
已有0條留言
- 還沒有人留言評論。精彩留言會獲得點贊!
1