誘導改善的免疫反應的疫苗的制作方法
【技術領域】
本發明涉及一種誘導改善的免疫反應的疫苗組合物。
背景技術:
最近,免疫佐劑備受矚目,并以多種方式用作宮頸癌疫苗及流感疫苗等。此類免疫佐劑中細菌DNA從1960年代開始,作為抗癌劑受到矚目,并直到現在,研究仍在繼續,但由于功效不足,無法單獨用作抗癌劑(Glick,J.L.The specificity of inhibition of tumor cell viability by DNA.CancerRes.27:2338,1967)。但眾所周知的是,細菌DNA無任何大的副作用,能夠對多種免疫細胞進行活性化,尤其,作為佐劑具有很多優點(McCluskie MJ,etal..CpG DNA is a potent enhancer of systemic and mucosal immune responses against hepatitis B surface antigen with intranasal administration to mice.JImmunol.Nov1;161(9):4463-6.1998)。
對細菌DNA的此類效果,日本的Yamamoto等主張包括CG的序列起到重要的作用,并通過美國Krieg等的研究廣為流傳(Yamamoto S.et al.Unique palindromic sequences in synthetic oligonucleotides are required to induce IFN and augment IFN-mediated natural killer activity.J.Immunol.148:4072,1992;Krieg AM,Antitumor applications of stimulating toll-like receptor 9with CpG oligodeoxynucleotides.Curr.Oncol.Rep.Mar;6(2):88-95.2004)。尤其在1990年代中期以后,通過CPG公司相關研究的結果,通過DNA抗癌劑確認了包含未甲基化的CG的合成DNA(CpG-ODN)的可能性,并研究了用于抑制合成DNA的分解的二酯鍵中的S置換等。通過此類努力,CpG-ODN相關產品不僅作為佐劑,還能作為抗癌劑進行臨床試驗(http://www.coleypharma.com)。
但是,對CpG-ODN的二酯鍵的S置換的免疫原性問題及仍舊不足的抗癌效果等依舊成為有待解決的問題。現在,臨床上所使用的CPG7909是引發anti-DNA Ab的硫代磷酸寡核苷酸(Clin Immunol.2001Aug;100(2):157-63),而這與系統性紅斑狼瘡(SLE,systemic lupus erythematosus)等自我免疫疾病具有密切的關系(J Clin Immunol.1986July;6(4):292-8)。并且,眾所周知,硫代磷酸結構起到TI Ag作用,混亂對感染的免疫防御功能(Mol Immunol.1998Dec;35(18):1161-70)。
從1950年代開始廣為流傳其抗癌效果的脂多糖(LPS)的情況下,僅僅由于ng水準的污染,也能引起基于敗血癥的死亡的毒性,而很難使用。尤其,一般的意見是,LPS和DNA的結合呈現嚴重的毒性,并且,在有關DNA的醫藥品中,LPS的清除被理解為非常重要的過程(Gao JJ.et.al,Bacterial DNA and lipopolysaccharide induce synergistic production of TNF-alpha through a post-transcriptional mechanism.JImmunol166(11):6855-60,2001)。在功效方面,基于LPS的免疫增加雖然比基于DNA的更加強大,但由于不是抗癌的重要的T楨1型,而是T楨2型,因此,認為并不適合用作抗癌劑(Lebman DA et al Interleukin 4causes isotype switching to IgE in T cell-stimulated clonal B cell cultures.J Exp Med.Sep 1;168(3):853-62.1988)。尤其,由于T楨2型的免疫活性抑制T楨1型的免疫活性,因此,LPS的此類特性更難使LPS用作抗癌劑(Rengarajan J et al.Transcriptional regulation of Th1/Th2polarization.Immunol Today.Oct;21(10):479-83.2000)。
對LPS的減毒試圖不斷地研究,尤其,能夠在通過聚糖鏈的清除或脂質A的脫酰基化來減少毒性方向獲得成功(Katz SS et al Deacylation of lipopolysaccharide in whole Escherichia coli during destruction by cellular and extracellular components of a rabbit peritoneal inflammatory exudate.J Biol Chem.Dec 17;274(51):36579-841999)。尤其,通過清除LPS聚糖鏈所獲得的脂質A的磷酸化來獲得的單磷酸脂質A(MPL,monophosphoryl lipid)的情況下,作為清除LPS的毒性的免疫抗癌劑來研發,但其效果微不足道(http://www.corixa.com)。
對此,本申請人已研發過一種補充上述多個免疫佐劑的缺點的新免疫佐劑(韓國專利授權第0740237號(2007.07.10))。
本說明書全文參照了多篇論文及專利文獻,并表示了其引用。所引用的論文及專利文獻的公開內容全部插入到本說明書中作為參照,從而更加明確地說明本發明所屬技術領域的水準及本發明的內容。
技術實現要素:
【發明要解決的技術課題】
本發明者為了研發一種能夠解決現有疫苗的問題,尤其,能夠增強疫苗的功效的新疫苗組合物而付出了努力。其結果,本發明者在利用多種病原體的抗原的免疫化過程中,作為免疫佐劑將脫酰基化非毒性脂寡糖(LOS,Lipooligosaccharide)用作免疫佐劑的情況下,已確認能夠誘導比現有的疫苗得到大大改善的免疫反應,成為用于預防多種疾病的優秀的疫苗,從而完成了本發明。
因此,本發明的目的在于,提供一種疫苗藥劑學組合物。
本發明的另一目的在于,提供一種樹突狀細胞的成熟化誘導用組合物。
本發明的其他目的及優點通過下述的發明內容,權利要求書及附圖將會變得更加明確。
【解決課題的技術方案】
根據本發明的一實施方式,本發明提供一種疫苗藥劑學組合物。該疫苗藥劑學組合物包含:
(a)病原體衍生抗原,其選自結核菌抗原、炭疽菌抗原、甲型肝炎病毒(HAV,Hepatitis A virus)的抗原、乙型肝炎病毒(HBV,Hepatitis B virus)的抗原、丙型肝炎病毒(HCV,Hepatitis C virus)的抗原、人類免疫缺陷病毒(HIV,human immunodeficiency virus)的抗原、流感病毒的抗原、單純性皰疹病毒(HSV,Herpes simplex virus)的抗原、B型流感嗜血桿菌(Hib,Haemophilus influenzae type b)的抗原、腦膜炎奈瑟菌(Neisseria meningitidis)的抗原、白喉棒狀桿菌(Corynebacterium diphtheria)的抗原、百日咳桿菌(Bordetella pertussis)的抗原、破傷風梭菌(Clostridium tetani)的抗原及水痘病毒(Varicella virus)的抗原組成的組中選擇;
(b)脫酰基化非毒性脂寡糖(LOS,Lipooligosaccharide);及
(c)藥劑學上接受的載體。
本發明者為了研發一種能夠解決現有疫苗的問題,尤其,能夠增強疫苗的功效的新疫苗組合物而付出了努力。其結果,本發明者在利用多種病原體的抗原的免疫化過程中,作為免疫佐劑將脫酰基化非毒性脂寡糖(LOS,Lipooligosaccharide)用作免疫佐劑的情況下,已確認能夠誘導比現有的疫苗得到大大改善的免疫反應,能夠成為用于預防多種疾病的優秀的疫苗。
本發明對引發結核的結核菌的疫苗有用。
本發明的結核疫苗中所利用的結核菌抗原包含以往公知的多種結核菌的衍生抗原,優選為選自65kD熱休克蛋白(HSP65)、抗原85A(Ag85A)、抗原85B、抗原85C、ESAT-6、Des蛋白、MPT32、MPT51、MPT63、MPT64、HspX及磷酸鹽結合蛋白1的組中的至少1種抗原。
更優選地,本發明的結核疫苗所利用的結核菌抗原是選自抗原85A(Ag85A)、ESAT-6、HspX及磷酸鹽結合蛋白1(Phosphate binding protein 1)的組中的至少1種抗原。
并且,本發明對引發炭疽病的炭疽疫苗有用。
本發明的炭疽疫苗中所利用的炭疽菌抗原包含以往公知的多種炭疽菌的衍生抗原,優選為保護性抗原(PA,protective antigen)、致死抗原(LF,lethal antigen)或水腫抗原(EA,edema antigen),更優選為保護性抗原(PA,protective antigen)。
并且,本發明對引發A型肝炎的HAV疫苗有用。本發明的HAV疫苗中所利用的HAV抗原包含以往公知的多種HAV衍生抗原,優選為選自活減毒HAV、惰性減毒HAV、VP1、VP2、VP3、VP4、2A、2B、2C、3A、3B、3C及3D蛋白的組中的抗原,更優選為惰性減毒HAV。
本發明對引發B型肝炎的HBV疫苗有用。本發明的HBV疫苗中所利用的HBV抗原包含以往公知的多種HBV衍生抗原,優選地,適合本發明的HBV抗原是HBV核心抗原(HBcAg,core antigen of HBV)或HBV表面抗原(HBsAg,surface antigen of HBV),更優選為HBV表面抗原(HBsAg,surface antigen of HBV)。
本發明對引發C型肝炎的HCV疫苗有用。本發明的HCV疫苗中所利用的HCV抗原包含以往公知的多種HCV衍生抗原,優選地,適合本發明的HCV抗原是E1、E2、NS3或NS3/4a抗原,更優選為NS3。
本發明對引發AIDS的HIV疫苗有用。本發明的HIV疫苗中所利用的HIV抗原包含以往公知的多種HIV衍生抗原,優選地,適合本發明的HIV抗原是Gag(p55gag)、Pol、Vif、Vpr、Tat、Rev、Vpu、Env或Nef抗原,更優選為Env抗原。
本發明對引發流感等的流感疫苗有用。本發明的流感疫苗中所利用的流感病毒的抗原包含以往公知的多種流感病毒衍生抗原,優選地,適合本發明的流感病毒的抗原是包膜糖蛋白HA或NA,更優選為HA。
本發明對引發單純皰疹的HSV疫苗有用。本發明的HSV疫苗中所利用的HSV的抗原包含以往公知的多種HSV衍生抗原,優選地,適合本發明的HSV抗原是糖蛋白gB、gD或gH,更優選為gB或gD。
本發明對引發髓膜炎等的Hib疫苗有用。本發明的Hib疫苗中所利用的Hib的抗原包含以往公知的多種Hib衍生抗原,優選地,適合本發明的Hib抗原是磷酸多核糖核醇(Haemophilus type b capsular polysaccharide(PRP,polyribosyl-ribitol-phosphate))或其共軛物,更優選為PRP和破傷風(tetanus)毒素的共軛物。
本發明對引發腦膜炎等的腦膜炎奈瑟菌(Neisseria meningitidis)疫苗有用。本發明的疫苗中所利用的腦膜炎奈瑟菌的抗原包含以往公知的多種腦膜炎奈瑟菌衍生抗原,優選地,適合本發明的腦膜炎奈瑟菌抗原是PI、PII、PIII、菌毛蛋白、脂多糖、鐵結合蛋白或蛋白酶體,更優選為蛋白酶體。
本發明對引發白喉等的白喉棒狀桿菌(Corynebacterium diphtheria)疫苗有用。本發明的疫苗中所利用的白喉棒狀桿菌的抗原包含以往公知的多種白喉棒狀桿菌衍生抗原,優選地,適合本發明的白喉棒狀桿菌的抗原是白喉毒素。
本發明對引發百日咳的百日咳桿菌(Bordetella pertussis)疫苗有用。本發明的疫苗中所利用的百日咳桿菌的抗原包含以往公知的多種百日咳桿菌衍生抗原,優選地,適合本發明的百日咳桿菌的抗原是百日咳毒素。
本發明對引發破傷風等的破傷風梭菌(Clostridium tetani)疫苗有用。本發明的疫苗中所利用的破傷風梭菌的抗原包含以往公知的多種破傷風梭菌衍生抗原,優選地,適合本發明的破傷風梭菌的抗原是破傷風毒素。
本發明對引發水痘等的水痘病毒(Varicella virus)疫苗有用。本發明的疫苗中所利用的水痘病毒的抗原包含以往公知的多種水痘病毒衍生抗原,優選地,適合本發明的水痘病毒的抗原是活減毒水痘病毒、惰性減毒水痘病毒、gpI或gpII,更優選為活減毒水痘病毒。
本發明的疫苗組合物中最大的特征在于,將非毒性脂寡糖(LOS,Lipooligosaccharide)作為免疫佐劑來利用。本說明書中首次采用的術語“非毒性脂寡糖(LOS,Lipooligosaccharide)”為脂多糖(LPS,lipopolysaccharide)的變形體,具有比天然(natural occurring)脂多糖更短的糖鏈,從而意味著分子量小。優選地,脫酰基化之前的脂寡糖的分子量為5000~10000Da。術語“脫酰基化脂寡糖”意味著在此類脂寡糖中清除與脂質A的葡萄糖胺通過-C(O)O-鍵相結合的脂肪酸,與LPO相比,毒性大大減少。脂肪酸在脂質A的葡萄糖胺通過-C(O)O-鍵及-C(O)NH-鍵相結合。本發明的脫酰基化脂寡糖表示通過脂質A的脫酰基化,來清除通過-C(O)O-鍵相結合的脂肪酸。
脫酰基化非毒性脂寡糖能夠通過多種方法來制備,但也能通過作為本發明者的現有專利的韓國專利授權第0456681號、WO2004/039413;韓國專利授權第0740237號及WO2006/121232中公開的方法來制備。例如,將強堿(2N NaOH)處理在脂多糖中進行脫酰基化,并從脂質A中清除一部分脂肪酸,來進行脫毒化。
根據本發明的優選實施例,在本發明中用作免疫佐劑的脫酰基化非毒性脂寡糖是對脂多糖進行堿處理,來實現脫酰基化而被非毒性化的。上述堿基的優選例包含NaOH、KOH、Ba(OH)2、CsOH、Sr(OH)2、Ca(OH)2、LiOH、RbOH及Mg(OH)2,更優選為NaOH、KOH、Ba(OH)2、Ca(OH)2、LiOH及Mg(OH)2,進而優選為NaOH、KOH及Mg(OH)2,最優選為NaOH。
脂多糖的毒性程度能夠通過所屬領域公知的多種方法來分析。例如,可對處理脂多糖的急性單核細胞細胞白血病(THP-1,Acute monocytic leukemia)分泌的腫瘤壞死因子-α(TNF-α,tumor necrosis factor-α)的量進行測定,來分析毒性。本發明的脫酰基化非毒性脂寡糖與現有的脂多糖相比,誘導相對少量的腫瘤壞死因子-α。
作為本發明中所利用的免疫佐劑,脫酰基化非毒性脂寡糖的又一特征在于,分子量比一般以往所利用的脂多糖要少。優選地,本發明中所利用的脫酰基化非毒性脂寡糖具有1500~10000Da的分子量,更優選為2000~5000Da,進而優選為2000~4000Da,尤其優選為3000~4000Da,最優選為3200~37000Da。此類分子量能夠通過所屬領域常規的方法來測定,例如,利用基質輔助激光解吸電離質譜(MALDI-MASS,matrix assisted laser desorption/ionization mass spectrometry)來測定。
根據本發明的優選實施例,作為本發明的免疫佐劑的脫酰基化非毒性脂寡糖來源于大腸桿菌(E.coli),最優選地,來源于作為本發明的單獨的菌的E.coli EG0021(KCCM 10374)。
本發明中所利用的脫酰基化非毒性脂寡糖在免疫促進(immunostimulatory)功效方面與現有的免疫佐劑相比更加優秀,與此同時,毒性也更加減少,因此,非常適合本發明的疫苗組合物。如下述實施例所論證一樣,比起為了清除脂多糖的毒素,通過清除脂多糖的聚糖鏈獲得的脂質A的磷酸化來獲得的單磷酸脂質A,本發明的脫酰基化非毒性脂多糖的毒性更少。
并且,如下述實施例所示,將本發明的脫酰基化非毒性脂多糖與多種病原體的抗原一同給藥的情況下,能夠大大增加免疫反應。
本發明的疫苗組合物僅僅通過其基本的組成,病原體的抗原和脫酰基化非毒性脂寡糖,也能誘導充分的免疫反應,從而發揮對特定疾病的預防功效。選擇性地,本發明的疫苗組合物還可包含其他免疫佐劑,例如,包含選自包含Mg、Ca、Sr、Ba及Ra的組中的第二族元素、選自包含Ti、Zr、Hf及Rf的組中的第四族元素,或鋁的鹽或其水合物。優選地,上述鹽與氧化物、過氧化物、氫氧化物、碳酸鹽、磷酸鹽、焦磷酸鹽、磷酸氫鹽、磷酸二氫、硫酸鹽或硅酸鹽一同形成。例如,能夠在本發明的疫苗組合物中追加利用的免疫佐劑包含氫氧化鎂、堿式碳酸鎂五水合、二氧化鈦、碳酸鈣、氧化鋇、氫氧化鋇、過氧化鋇、硫酸鋇、硫酸鈣、焦磷酸鈣、碳酸鎂、氧化鎂、氫氧化鋁、磷酸鋁、十二水合硫酸鋁鉀(Alum)。最優選地,在本發明的疫苗組合物中,能夠追加利用的免疫佐劑為氫氧化鋁。
包含于本發明的疫苗藥劑學組合物的、在藥劑學上接受的載體是在制劑時通常利用的,其包含乳糖、右旋糖、蔗糖、山梨糖醇、甘露糖醇、淀粉、阿拉伯樹膠、磷酸鈣、藻酸鹽、明膠、硅酸鈣、微晶纖維素、聚乙烯吡咯烷酮、纖維素、水、糖漿、甲基纖維素、羥基苯甲酸甲酯、羥基苯甲酸丙酯、滑石、硬脂酸鎂及礦物油,但并不局限于此。本發明的藥劑學組合物除了上述成分之外,還可包含潤滑劑、潤濕劑、增甜劑、香味劑、乳化劑、懸浮劑、保存劑等。藥劑學上接受的適合的載體及制劑在Remington's Pharmaceutical Sciences(19th ed.,1995)中有詳細記載。
本發明的藥劑學組合物能夠以口服或非口服方式進行給藥,優選為非口服給藥,就非口服給藥而言,能夠以靜脈內注射、皮下注射、肌肉注射、腹腔注射、經皮等方式進行給藥。
本發明的藥劑學組合物的適合的給藥劑量會根據制劑方法、給藥方式、患者的年齡、體重、性別、病態、飲食、給藥時間、給藥途徑、排泄速度及反應靈敏性之類的因素,能夠進行多種處方。另一方面,本發明的藥劑學組合物的口服給藥劑量,按每日優選為0.0001~1000mg/kg(體重)。
本發明的藥劑學組合物根據所屬領域技術人員能夠容易實施的方法,利用藥劑學上接受的載體和/或賦形劑進行制劑化,從而以單位容量形態進行制備或內置于多容量容器來制備。此時,劑型可以是油性或水性介質中的溶液、懸浮液或乳化液形態,也可以是浸膏劑、粉末劑、顆粒劑、片劑或膠囊劑,并且,還可包含分散劑或穩定劑。
本發明的疫苗組合物大大誘導對病原體的免疫反應,從而對特定疾病具有優秀的預防功效之外,由于用作免疫佐劑的脫酰基化非毒性脂寡糖幾乎沒有毒性,因此,在穩定性方面也非常優秀。
本發明所利用的脫酰基化非毒性脂寡糖在多種病原體抗原中,對結核菌抗原、炭疽菌抗原、HAV抗原、HBV抗原、HCV抗原、HIV抗原、流感病毒的抗原、HSV抗原、Hib(Haemophilus influenzae type b)的抗原、腦膜炎奈瑟菌抗原、白喉棒狀桿菌、百日咳桿菌抗原、破傷風梭菌抗原及水痘疾病抗原起著尤其促進優秀的免疫反應的作用。脫酰基化非毒性脂寡糖的此類優秀的免疫反應促進效果在下述的實施例中有論證。
根據本發明的另一實施方式,本發明提供一種包含脫酰基化非毒性脂寡糖的樹突狀細胞的成熟化誘導用組合物。
根據本發明的又一實施方式,本發明提供一種包括將脫酰基化非毒性脂寡糖處理在未成熟樹脂細胞的成熟化樹突狀細胞的制備方法。
在制備樹突狀細胞疫苗時,應對從生物體中分離的樹突狀細胞進行成熟化。在此過程中,能夠非常有用地利用脫酰基化非毒性脂寡糖。
根據樹突狀細胞疫苗的制備方法,未成熟的樹突狀細胞來源于單核細胞或造血干細胞,更優選地,來源于單核細胞。從單核細胞中誘導樹突狀細胞的方法更加優選的理由在于,與使用多功能性造血干細胞(pluripotent hematopoietic stem/progenitor cells)的方法相比,能夠在較短的步驟內獲得均等性質的樹突狀細胞。根據本發明的優選實施例,上述單核細胞是從外周血中獲得的。從外周血內的祖細胞(progenitor cell),例如,單核細胞或造血干細胞)中獲得未成熟的樹突狀細胞的方法能夠通過從外周血內其他細胞中分離上述祖細胞,來進行樹突狀細胞進行分化和誘導的過程,并且,還能進行以未從外周血內其他細胞中分離的狀態,將上述祖細胞分化和誘導為樹突狀細胞的過程。
之后,將從外周血中獲得的單核細胞分化和誘導為樹突狀細胞。為了分化為樹突狀細胞,在包含適合的細胞因子的培養基內培養單核細胞。所利用的細胞因子是粒細胞-巨噬細胞集落促進因子(GM-CSF,Granulocyte macrophage colony stimulating factor)、白細胞介素-4(IL-4,interleukin-4),白細胞介素-13(IL-13)或它們的組合。所添加的細胞因子的適合的量是充分分化為樹突狀細胞的量,因此,是各研究人員在實驗室中以經驗進行選擇而優選的量。
從單核細胞中分化樹突狀細胞為目的的第一次培養時間,在使用細胞因子來進行的情況下,通常在5天以上,優選為6~7天。
利用在上述的分化過程中獲得的未成熟的樹突狀細胞,最終實施成熟化過程。即,在包含本發明的脫酰基化非毒性脂寡糖的具有培養液的培養基中進行培養,來將樹突狀細胞進行成熟化。第二次培養的時間通常在10小時以上,優選為20小時以上,例如,1~3天。
本發明能夠有效地制備成熟化的樹突狀細胞疫苗。
【發明效果】
歸納本發明的特征及優點如下。
(a)本發明的疫苗組合物利用脫酰基化非毒性脂寡糖作為病原體衍生抗原和免疫佐劑。
(b)作為免疫佐劑的脫酰基化非毒性脂寡糖對特定病原體的免疫反應中發揮非常優秀的免疫促進功效的同時,幾乎沒有毒性,因此,穩定性非常優秀。
(c)并且,脫酰基化非毒性脂寡糖能夠有效地制備成熟化的樹突狀細胞疫苗。
【附圖說明】
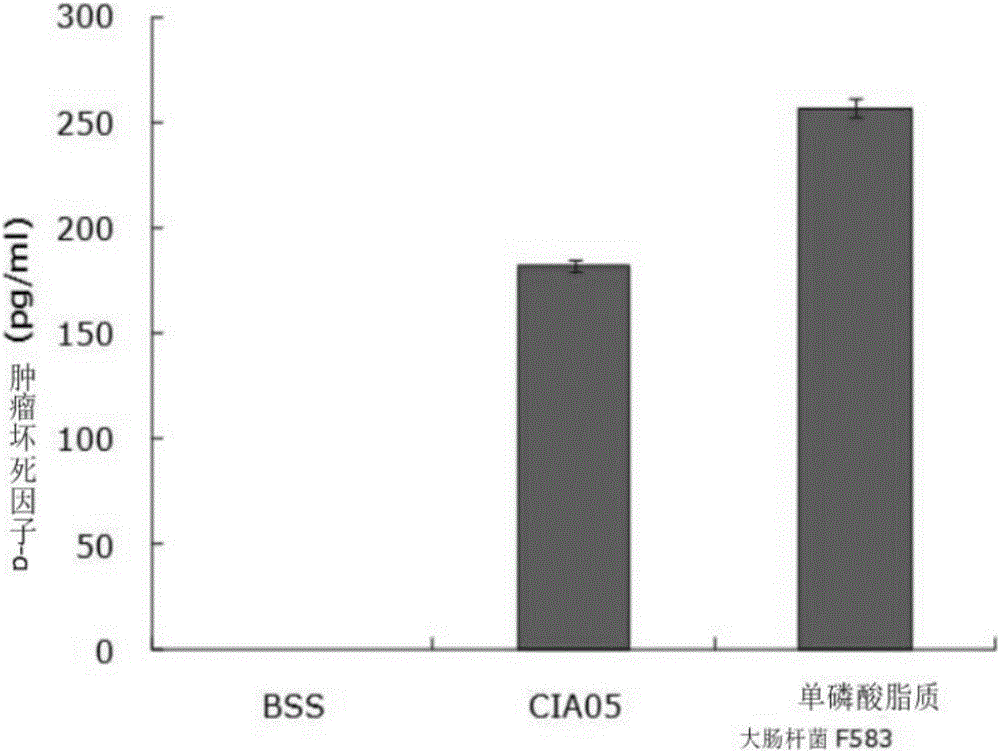
圖1是表示新免疫佐劑(CIA05)的細胞毒性(分泌腫瘤壞死因子-α)比單磷酸脂質A更低的圖表。
圖2a~圖2d是對本發明的結核疫苗的功效進行分析的實驗結果。圖2a~圖2d中,作為針對結核菌的抗原,利用Ag85A、HspX、磷酸鹽結合蛋白1(Phosphate binding protein 1)及ESAT-6。
圖2e是對Ag85A、HspX、ESAT-6及38-kDa的免疫促進功能進行比較的實驗結果。圖2e中利用了CIA05 0.5μg及各個抗原2μg。
圖3是對本發明的炭疽疫苗的功效進行分析的實驗結果。
圖4是對本發明的A型肝炎疫苗的功效進行分析的實驗結果。
圖5是對本發明的B型肝炎疫苗的功效進行分析的實驗結果。
圖6是對本發明的C型肝炎疫苗的功效進行分析的實驗結果。
圖7是對本發明的HIV疫苗的功效進行分析的實驗結果。
圖8是對本發明的流感疫苗的功效進行分析的實驗結果。
圖9a~圖9b是對本發明的HSV-2(Herpes simplex virus type 2)疫苗的功效進行分析的實驗結果。圖9a及圖9b中,作為各個抗原,利用了gD抗原及gB抗原。
圖9c是對gD抗原及gB抗原的免疫促進功能進行比較的實驗結果。圖9c中利用了CIA05 0.5μg及各個抗原2μg。
圖10是對本發明的Hib(Haemophilus influenzae type b)疫苗的功效進行分析的實驗結果。
圖11是對本發明的腦膜炎奈瑟菌疫苗的功效進行分析的實驗結果。
圖12a~圖12c是對本發明的DPT疫苗的功效進行分析的實驗結果。圖12a~圖12c中,作為各個抗原,分別利用了白喉毒素、百日咳毒素及破傷風毒素。
圖12d是對白喉毒素、百日咳毒素及破傷風毒素的免疫促進功能進行比較的實驗結果。圖12d中,利用了CIA05 0.5μg及各個抗原2μg。
圖13是對本發明的水痘疫苗疫苗的功效進行分析的實驗結果。
圖14a是在小鼠骨髓來源的樹突狀細胞(BMDCs,Bone marrow-derived dendritic cells)中,分析對樹突狀細胞的成熟化的CIA05(實施例1中制備的免疫佐劑)的功效的結果。
圖14b是在人單核細胞來源的樹突狀細胞(MDDCs,Monocyte-derived dendritic cells)中,分析對樹突狀細胞的成熟化的CIA05(實施例1中制備的免疫佐劑)的功效的結果。
【實施方式】
以下,通過實施例對本發明進行更加詳細的說明。這些實施例僅僅是為了對本發明進行更加詳細的說明,根據本發明的主旨,本發明請求保護的范圍并不局限于這些實施例,這對于本發明所屬技術領域的普通技術人員來說是顯而易見的。
【實施例】
【實施例1:作為新免疫佐劑的非常短的脂寡糖(LOS)CIA05的生產】
本發明者已經從生存在健康的人的腸道中的大腸桿菌中發掘了具有脂寡糖的糖鏈的長度非常短的脂多糖的菌株(E.coli EG0021),并于2002年5月2日,將此菌株寄托于寄托機構—韓國微生物保管中心,并獲得了寄托編號KCCM10374(參照:韓國專利授權第0456681號;WO2004/039413;韓國專利授權第0740237號;WO2006/121232)。從上述菌株的脂多糖的純化是根據韓國專利授權第0456681號;WO2004/039413;韓國專利授權第0740237號;WO2006/121232中公開的方法來實施的。利用MALDI-MASS(Shimadz公司,Axima-LNR V 2.3.5(Mode Liner,Power:106))來測定純化的脂多糖的分子量,測定結果,確認了具有約3千5百Da的分子量。純化的脂多糖根據韓國專利授權第0456681號;WO2004/039413;韓國專利授權第0740237號;WO2006/121232中公開的方法,根據公開的實驗方案進行了脫毒化。簡單地,在3mg/ml的上述純化的脂多糖中,以1:1的量混合2N NaOH后,在60℃下每隔10分鐘振蕩一次,如此進行140分鐘脫酰基化,投入初期0.2N NaOH量的約1/5左右的1N乙酸,來滴定至pH7.0。在pH滴定后,進行乙醇沉淀,從而獲得非毒性脂寡糖(CIA05)。
【實施例2:新免疫佐劑CIA05和現有免疫佐劑單磷酸脂質A之間的毒性比較實驗】
對本發明的疫苗中用作免疫佐劑的上述實施例中所制備的CIA05和現有的宮頸癌疫苗中所使用的單磷酸脂質A的毒性進行了比較分析。接收健康的人的血液,來分離外周血單核細胞(PBMC,Peripheral Blood Mononuclear Cell),在24孔培養皿中以5×105cell/ml的濃度培養上述外周血單核細胞。此時,使用在RPMI1640(Gibco公司)中混合10%的胎牛血清(FBS,fetal calf serum;Gibco公司)而得的培養液,而每個孔的容量均分為1ml。以如下條件分別處理了準備好的培養皿。1)陰性對照組:平衡鹽溶液(BBS,Balanced salt solution)100μl;2)脫酰基化非毒性脂寡糖(CIA05)10μg/100μl:3)單磷酸脂質A(E.coli F583MPL)10μg/100μl。
經過12小時之后,收集已處理的培養液進行離心分離,并利用酶聯免疫吸附測定方法(ELISA,enzyme linked immunosorbent assay),使用酶聯免疫吸附測定試劑盒(R&D system公司,DY210)對急性單核細胞白血病(THP-1,Acute monocytic leukemia)中分泌的作為炎性蛋白的腫瘤壞死因子-α的量分別進行測定。實驗結果表明,如圖1所示,毒性比以往用作疫苗佐劑的單磷酸脂質A,大約少1/3左右。
【實施例3:本發明的結核疫苗的功效分析】
【結核疫苗抗原和免疫佐劑CIAO5的免疫】
為了在結核疫苗中確認免疫佐劑CIAO5的功效,使用了結核菌株(Mycobacterium tuberculosis)的4種抗原。這4種抗原為Ag85A(32-kDa)、HspX(16-kDa)、Phosphate binding protein 1(38-kDa)及ESAT-6(6-kDa)。分別混合每2μg的上述4種抗原,每隔一周向6周齡的Balb/c小鼠(SLC,日本)注射3次。此時,單獨注射抗原或者包含Alum(氫氧化鋁;Brenntag公司,德國)100μg或CIA050.5μg,以最終體積為100μl來進行注射。
【結核疫苗抗原的特意血清抗體的滴定】
為了測定免疫后的血清內結核抗原的特意抗體滴度,使用了終點稀釋酶聯免疫吸附測定(ELSIA,Enzyme-Linked Immunosorbent Assay)方法。以1μg/ml的濃度向96孔板分別涂敷(4℃,一夜)100μl的Ag85A(32-kDa)、HspX(16-kDa)、Phosphate binding protein1(38-kDa)及ESAT-6(6-kDa)后,利用1%牛血清白蛋白(BSA,bovine serum albumin)300μg來封閉(常溫,1小時)。封閉之后,利用包含0.05%Tween-20的磷酸鹽緩沖溶液(PBS,Phosphate-Buffered Saline)清洗3次,并將免疫后所獲得的血清100μl進行反應(37℃,2小時)。為了確認結合抗原的特意抗體,將辣根過氧化物酶-共軛物抗-小鼠IgG(Zymed)進行反應后,添加四甲基聯苯胺(TMB,tetramethylbenzidine,BD生物科學公司,55555214),并利用1N H2SO4終止反應。實驗結果,在450nm測定吸光度,從而確認了免疫后回收的血清內結核抗原的特意抗體的量。
【結核疫苗中的免疫佐劑CIA05的功效分析】
對結核菌株的4種抗原和免疫佐劑CIA05的功效進行分析的結果,在Ag85A(32-kDa)抗原的情況下,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum100μg)左右增加的結核抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約128倍(CIA05 1μg)左右增加的結核抗原-特意抗體的生成(圖2a)。并且,在利用CIA05的情況下,結核抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對結核抗原Ag85A是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)
在HspX(16-kDa)抗原中,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的結核抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約128倍(CIA05 1μg)左右增加的結核抗原-特意抗體的生成(圖2b)。并且,在利用CIA05的情況下,結核抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對結核抗原HspX(16-kDa)是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。
對于Phosphate binding protein 1(38-kDa)抗原而言,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的結核抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約8倍(CIA05 0.5μg)或大約64倍(CIA05 1μg)左右增加的結核抗原-特意抗體的生成(圖2c)。并且,在利用CIA05的情況下,結核抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對結核抗原Phosphate binding protein 1(38-kDa)是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。
對于ESAT-6(6-kDa)抗原而言,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum100μg)左右增加的結核抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約128倍(CIA05 1μg)左右增加的結核抗原-特意抗體的生成(圖2d)。并且,在利用CIA05的情況下,結核抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對結核抗原ESAT-6(6-kDa)是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。
在結核抗原中,實施了一種用于選擇免疫促進功能最優秀的抗原的分析(圖2e)。實驗結果表明,在結核菌抗原中,依次以Ag85A、HspX、ESAT-6及38-kDa的順序具有免疫促進功能,由此能夠了解與本發明的CIA05最適合的抗原為Ag85A。
綜上所述,能夠了解作為免疫佐劑包含CIA05的本發明的結核疫苗在免疫功效,即疫苗功效方面非常優秀,并且,與作為現有的免疫佐劑的Alum相比,對結核抗原具有更加優秀的免疫促進功能。
【實施例4:本發明的炭疽疫苗的功效分析】
【作為炭疽疫苗抗原的PA與免疫佐劑CIA05的免疫】
炭疽菌(Bacillus anthracis)以3種蛋白生成2種毒素,在此,3種蛋白由保護性抗原(PA,protective antigen)、致死抗原(LF,lethal antigen)及水腫抗原(EA,edema antigen)組成。其中,PA用于炭疽疫苗,并在本實驗中使用了重組全長PA抗原。以腹腔注射方式,每隔兩周向6周齡的雄性Balb/c小鼠3次注射10μg炭疽菌的PA抗原。使用10μg的PA抗原,使用0.5μg、1μg的CIA05,使用50μg、100μg的Alum,使最終體積達到100μl。對陰性對照組每次注射100μl的磷酸鹽緩沖溶液(PBS,Phosphate-Buffered Saline,pH 7.3)。向小鼠注射3次之后,從小鼠身上采集全血,并通過離心分離獲得血清。為了測定免疫后血清內PA特意抗體的滴度,如實施例3一樣,使用終點酶聯免疫吸附測定(ELSIA)方法。在免疫化后,對血清內PA特意抗原進行分析的結果表明,與單獨免疫化PA抗原時相比,將0.5μg的CIA05作為免疫佐劑進行給藥時,增加10倍的PA特意抗原,而將1μg的CIA05作為免疫佐劑進行給藥時,增加21倍的PA特意抗原(圖3)。這表示CIA05與作為炭疽抗原的PA一同在炭疽疫苗中起到免疫佐劑的作用。并且,能夠確認混合使用CIA05和Alum(aluminum hydroxide),來用作免疫佐劑時,能夠增進PA特意免疫反應(圖3)。
綜上所述,作為免疫佐劑包含CIA05的本發明的炭疽疫苗,尤其是利用PA抗原和CIA05組合的炭疽疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例5:本發明的A型肝炎(Hepatitis A virus;HAV)疫苗的功效分析】
作為抗原,使用了MRC-5細胞株中培養的惰性HAV。每隔一周向6周齡的Balb/c小鼠注射3次惰性HAV。此時,單獨注射HAV或者包含CIA05進行注射。在注射3次過一周后,如同實施例3一樣分離血清,通過終點酶聯免疫吸附測定方法測定HAV特意抗體。
對于HAV抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約2倍(Alum 50μg)或大約4倍(Alum 100μg)左右增加的HAV抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約8倍(CIA05 0.5μg)或大約32倍(CIA05 1μg)左右增加的HAV抗原-特意抗體的生成(圖4)。并且,在利用CIA05的情況下,HAV抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對HAV抗原(惰性HAV)是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的HAV疫苗,尤其是利用惰性HAV和CIA05組合的HAV疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例6:本發明的B型肝炎(Hepatitis B virus;HBV)疫苗的功效分析】
作為HBV預防疫苗中的抗原,將作為HBV的表面抗原的HBsAg(adr subtype)在多形漢遜酵母(Hansenula polymorpha)中表達、純化并使其重組來使用。每隔一周向Balb/c小鼠注射3次作為HBV抗原的HBsAg 2μg。此時,單獨注射抗原或者包含Alum或CIA05進行注射。在注射3次過一周后,如同實施例3一樣分離血清,通過終點酶聯免疫吸附測定方法測定HBsAg特意抗體。
對于HBsAg抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約2倍(Alum 50μg)或大約4倍(Alum 100μg)左右增加的HBsAg抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約8倍(CIA05 0.5μg)或大約16倍(CIA05 1μg)左右增加的HBsAg抗原-特意抗體的生成(圖5)。并且,在利用CIA05的情況下,HBsAg抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對HBsAg抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的HBV疫苗,尤其是利用HBsAg和CIA05組合的HBV疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例7:本發明的C型肝炎(Hepatitis C virus;HCV)疫苗的功效分析】
為了在HCV疫苗中確認免疫佐劑CIA05的功效,購買HCV的重組NS3(Z00042,金斯瑞生物科技)作為抗原來使用。
每隔一周向6周齡Balb/c小鼠注射3次2μg的NS3抗原,單獨注射抗原或者包含Alum或CIA05進行注射。在注射3次過一周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定NS3特意抗體。
對于NC3抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約2倍(Alum 50μg)或大約4倍(Alum 100μg)左右增加的NC3抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約4倍(CIA05 0.5μg)或大約16倍(CIA05 1μg)左右增加的NC3抗原-特意抗體的生成(圖6)。并且,利用CIA05的情況下,NC3抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對NC3抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約4倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的HCV疫苗,尤其是利用NC3抗原和CIA05組合的HCV疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例8:本發明的HIV疫苗的功效分析】
為了在HIV疫苗中確認免疫佐劑CIA05的功效,購買HIV-1包膜(Env)抗原(H9909,西格馬公司)作為抗原來使用。每隔一周向6周齡Balb/c小鼠注射3次2μg的HIV-1包膜抗原,單獨注射抗原或者包含Alum或CIA05進行注射。在注射3次過一周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定HIV-1包膜抗原特意抗體。
對于HIV-1包膜抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的HIV-1包膜抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約64倍(CIA05 1μg)左右增加的HIV-1包膜抗原-特意抗體的生成(圖7)。并且,在利用CIA05的情況下,HIV-1包膜抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對HIV-1包膜抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的HIV疫苗,尤其是利用HIV-1包膜抗原和CIA05組合的HIV疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例9:本發明的流感疫苗的功效分析】
為了在流感疫苗中確認免疫佐劑CIA05的功效,使用了從3種流感病毒菌株中獲得的HA亞單位(subunit)(CJ公司,仁川,韓國)。3種流感病毒菌株為A/New Caledonia/20/99(HIN1)、A/Panama/2007/99(H3N2)及B/Shangdong/7/97。每隔四周向6周齡Balb/c小鼠注射2次來源于上述3種流感病毒菌株的HA subunit1.5μg。此時,單獨注射抗原或者包含25μg的Alum或25μg的CIA05進行注射。在注射2次過4周后,如同實施例1一樣,從小鼠身上分離血清,通過終點酶聯免疫吸附測定方法測定HA subunit特意抗體。
對于流感HA subunit抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約2倍(Alum 50μg)或大約4倍(Alum 100μg)左右增加的流感HA subunit抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約4倍(CIA05 0.5μg)或大約8倍(CIA05 1μg)左右增加的流感HA subunit抗原-特意抗體的生成(圖8)。并且,在利用CIA05的情況下,流感HA subunit抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對流感HA subunit抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約8倍(Alum100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約8倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的流感疫苗,尤其是利用流感HA subunit抗原和CIA05組合的流感疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例10:本發明的Ⅱ型單純皰疹病毒(HSV-2,Herpes simplex virus type 2)疫苗的功效分析】
為了在HSV-2疫苗中確認免疫佐劑CIA05的功效,將HSV-2的糖蛋白gD和gB作為抗原來使用。由于2種抗原的糖化非常重要,因此,為了準確的糖化,在作為動物細胞的中國倉鼠卵巢(CHO,Chinese hamster ovary)細胞株中進行表達及純化(Boucher et al.,Detection of antibodies to herpes simplex virus type 2with a mammalian cell line expressing glycoprotein gG-2.Clin.Diagn.Virol.1(1):29-38(1993))。每隔一周向6周齡Balb/c小鼠注射3次在CHO細胞株中進行表達、純化的2μg的2種抗原,而實驗組單獨注射抗原或者包含Alum或CIA05進行注射。在注射3次過1周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定作為HSV-2抗體的gD和gB特意抗體。
對于gD抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約8倍(Alum 50μg)或大約32倍(Alum 100μg)左右增加的gD抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約24倍(CIA05 0.5μg)或大約128倍(CIA05 1μg)左右增加的gD抗原-特意抗體的生成(圖9a)。并且,在利用CIA05的情況下,gD抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對gD抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約16倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約2倍(Alum 50μg)或大約16倍(Alum 100μg)。
對于gB抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約8倍(Alum 50μg)或大約32倍(Alum 100μg)左右增加的gB抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約32倍(CIA051μg)左右增加的gB抗原-特意抗體的生成(圖9b)。并且,利用CIA05的情況下,gB抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對gB抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約16倍(Alum100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約16倍(Alum 100μg)。
HSV-2抗原中,實施了一種用于選擇免疫促進功能最優秀的抗原的分析(圖9c)。實驗結果表明,在HSV-2抗原中,gD抗原的免疫促進功能比gB抗原優秀,由此,能夠了解與本發明的CIA05最適合的抗原為gD抗原。
綜上所述,作為免疫佐劑包含CIA05的本發明的HSV-2疫苗,尤其是利用gD抗原和CIA05組合的HSV-2疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例11:本發明的b型流感嗜血桿菌(Hib,Haemophilus influenzae type b)疫苗的功效分析】
作為Hib抗原,購買乙型流感嗜血桿菌疫苗安爾寶(ActHIB)(b型流感嗜血桿菌(破傷風類毒素,賽諾菲安萬特公司)來使用。每隔兩周向Balb/c小鼠注射2次ActHIB。此時,單獨注射疫苗或者包含Alum或CIA05進行注射。在注射2次過1周后,如同實施例1一樣,從已注射的小鼠身上分離血清,通過終點酶聯免疫吸附測定方法測定Hib特意抗體。
對于ActHIB抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約16倍(Alum 100μg)左右增加的ActHIB抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA05 0.5μg)或大約32倍(CIA05 1μg)左右增加的ActHIB抗原-特意抗體的生成(圖10)。并且,在利用CIA05的情況下,ActHIB抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對ActHIB是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約8倍(Alum50μg)或大約16倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約24倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的Hib疫苗,尤其是利用ActHIB和CIA05組合的Hib疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例12:本發明的腦膜炎奈瑟菌疫苗的功效分析】
為了在腦膜炎奈瑟菌疫苗中確認免疫佐劑CIA05的功效,從減毒的腦膜炎奈瑟菌中分離蛋白酶體,將其作為抗原來使用(Lowell et al.,Proteosome-lipopeptide vaccines:enhancement of immunogenicity for malaria CS peptides.Science,240:800-802(1980))。每隔兩周向6周齡的Balb/c小鼠注射2次抗原,實驗組組成單獨使用疫苗或者包含CIA05的2種實驗組。在注射2次過1周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定用作抗原的腦膜炎奈瑟菌蛋白酶體(Neisseria meningitidis proteosome)特意抗體。
對于腦膜炎奈瑟菌蛋白酶體抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約8倍(CIA05 0.5μg)或大約16倍(CIA05 1μg)左右增加的抗原-特意抗體的生成(圖11)。并且,在利用CIA05的情況下,抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對腦膜炎奈瑟菌蛋白酶體抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約16倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約16倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的腦膜炎奈瑟菌疫苗,尤其是利用腦膜炎奈瑟菌蛋白酶體和CIA05組合的腦膜炎奈瑟菌疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例13:本發明的白喉、破傷風、百日咳(DPT)疫苗的功效分析】
DPT是預防由白喉,百日咳,破傷風引起的疾病的疫苗。為了對此類疾病的疫苗確認CIA05的免疫佐劑的功效,購買來源于白喉棒狀桿菌的白喉毒素(DO564,西格瑪公司)作為白喉來使用,購買來源于百日咳桿菌的百日咳毒素(P7208,西格瑪公司)作為百日咳來使用,購買來源于破傷風梭菌的破傷風毒素(T3194,西格瑪公司)作為破傷風來使用。混合每2μg的3種毒素,每隔兩周向6周齡Balb/c小鼠注射2次。此時,單獨注射抗原或者包含Alum或CIA05進行注射。在注射2次過一周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定對于3種毒素的特意抗體。
對于白喉病毒抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約32倍(Alum 50μg)或大約64倍(Alum 100μg)左右增加的抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約32倍(CIA05 0.5μg)或大約128倍(CIA05 1μg)左右增加的抗原-特意抗體的生成(圖12a)。并且,在利用CIA05的情況下,抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對白喉毒素抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約8倍(Alum 50μg)或大約16倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。
對于百日咳病毒抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約16倍(Alum 100μg)左右增加的抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA050.5μg)或大約24倍(CIA051μg)左右增加的抗原-特意抗體的生成(圖12b)。并且,在利用CIA05的情況下,抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對百日咳毒素抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約32倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約8倍(Alum 50μg)或大約32倍(Alum 100μg)。
對于破傷風毒素抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA050.5μg)或大約32倍(CIA051μg)左右增加的抗原-特意抗體的生成(圖12b)。并且,在利用CIA05的情況下,抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對白喉毒素抗原是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。
在DPT抗原中,實施了一種用于選擇免疫促進功能最優秀的抗原的分析(圖12d)。實驗結果表明,免疫促進功能依次以白喉毒素、百日咳毒素及破傷風毒素的順序優秀,由此能夠了解與本發明的CIA05最適合的抗原為白喉毒素。
綜上所述,作為免疫佐劑包含CIA05的本發明的DPT,尤其是利用白喉毒素和CIA05組合的DPT疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例14:本發明的水痘疫苗功效分析】
為了在水痘疫苗中確認免疫佐劑CIA05的功效,作為抗原,使用了1VIA水痘疫苗株(1VIA Suduvax注射液,綠十字)。水痘疫苗株是一種減毒活疫苗形態的水痘疫苗。每隔兩周向6周齡的Balb/c小鼠注射2次水痘疫苗。此時,單獨注射疫苗或者包含CIA05進行注射。在注射2次過一周后,如同實施例1一樣分離血清,通過終點酶聯免疫吸附測定方法測定水痘菌株特意抗體。
對于水痘菌株抗原,利用Alum的實驗組與單獨使用抗原的實驗組相比,增進大約4倍(Alum 50μg)或大約8倍(Alum 100μg)左右增加的抗原-特意抗體的生成,在利用CIA05的情況下,與單獨使用抗原的實驗組相比,增進大約16倍(CIA050.5μg)或大約64倍(CIA051μg)左右增加的抗原-特意抗體的生成(圖13)。并且,在利用CIA05的情況下,抗原-特意抗體產量對給藥劑量依賴性地劇增,從而能夠了解CIA05對水痘菌株抗原(減毒的水痘菌株)是非常適合的免疫佐劑。一方面,作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用0.5μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約8倍(Alum 100μg)。作為免疫佐劑,將CIA05和Alum同時使用的情況下,比起只使用1.0μg的CIA05的情況,抗體產量增加大約4倍(Alum 50μg)或大約32倍(Alum 100μg)。
綜上所述,作為免疫佐劑包含CIA05的本發明的水痘疫苗,尤其是利用減毒的水痘菌株和CIA05組合的水痘疫苗在免疫功效,即疫苗功效方面非常優秀。
【實施例15:對樹突狀細胞的成熟化的CIA05的功效分析】
【人單核細胞衍生的DC細胞(MDDCs,Monocyte-derived DC cells)和小鼠骨髓樹突狀細胞(BMDC,Bone marrow DCs)的分離】
從健康的人身上采集約80ml的血液后,通過聚蔗糖泛影鈉TM梯度(Ficoll-PaqueTMgradient)分離法分離外周血單核細胞(PBMCs)。單核細胞(monocyte)是從外周血單核細胞(PBMCs)中利用anti-CD14微珠(microbeads)(免疫磁珠分離系統:MACS system)來分離的,并且,在24孔板中,以1×108cells/well的濃度,利用包含10%的FBS、IL-4、GM-CSF的RPMI1640培養基進行稀釋和培養,從而獲得人類單核細胞衍生的DC細胞。骨髓樹突狀細胞是在雌性BALB/c小鼠中,將骨髓細胞(bone marrow cell)在包含IL-4、GM-CSF的培養基中培養6天,并在第7天利用抗-CD11c-包被磁珠(anti-CD11c-coated magnetic beads)分離CD11c+cell。
【基于免疫佐劑CIA05的樹狀突細胞(DCs,Dendritic cells)表面標記表達增加分析】
通過上述方法分離人單核細胞衍生的DC細胞和小鼠的骨髓樹突狀細胞后,將脂多糖、單磷酸脂質A、CIA05分別以1μg/ml的濃度進行處理,在37℃下進行24小時反應。進行反應之后,在小鼠骨髓樹突狀細胞的表面確認了CD40、CD80及CD86,在人單核細胞衍生的DC細胞中分別通過流式細胞儀(flow cytometry)確認了HLA-DR、CD80及CD86。能夠確認的是,小鼠的骨髓樹突狀細胞的情況下,脂多糖、單磷酸脂質A及CIA05幾乎都類似地分布了作為DCs表面標記的CD40、CD80及CD86(圖14a)。但是,在人單核細胞衍生的DC細胞中能夠確認,與現在廣泛使用于疫苗免疫佐劑的單磷酸脂質A相比,作為單核細胞衍生的DC細胞的表面標記的HLA-DR、CD80及CD86在CIA05中有著更多的表達(圖14b)。這表示單磷酸脂質A在小鼠身上會維持脂多糖水準的功效,但在人身上會限制脂多糖水準的功效,并且,CIA05能夠解決此類問題。
綜上所述,上述實驗結果表明,免疫佐劑CIA05在多種疫苗中起著大大增加免疫功效,即疫苗功效的作用,尤其,對人類樹突狀細胞的成熟化也有顯著效果。
本說明書還包括下列內容:
1.一種疫苗藥劑學組合物,其特征在于,包含:
(a)病原體衍生抗原,其選自包含結核菌抗原、炭疽菌抗原、HAV的抗原、HBV的抗原、HCV的抗原、HIV的抗原、流感病毒的抗原、HSV的抗原、Hib的抗原、腦膜炎奈瑟菌的抗原、白喉棒狀桿菌的抗原、百日咳桿菌的抗原、破傷風梭菌的抗原及水痘病毒的抗原的組中;
(b)脫酰基化非毒性脂寡糖;以及
(c)藥劑學上接受的載體。
2.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述結核菌抗原是選自包含65kD熱休克蛋白、抗原85A、抗原85B、抗原85C、ESAT-6、Des蛋白、MPT32、MPT51、MPT63、MPT64、HspX及磷酸鹽結合蛋白1的組中的至少1種抗原。
3.根據實施方式2所述的疫苗藥劑學組合物,其特征在于,上述結核菌抗原是選自包含抗原85A、ESAT-6、HspX及磷酸鹽結合蛋白1的組中的至少1種抗原。
4.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述炭疽病抗原為保護性抗原、致死抗原或水腫抗原。
5.根據實施方式3所述的疫苗藥劑學組合物,其特征在于,上述炭疽菌抗原為保護性抗原。
6.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述HAV的抗原是選自包含活減毒HAV、惰性減毒HAV、VP1、VP2、VP3、VP4、2A、2B、2C、3A、3B、3C及3D蛋白的組中的抗原。
7.根據實施方式6所述的疫苗藥劑學組合物,其特征在于,上述HAV的抗原為惰性減毒HAV。
8.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述HBV的抗原為HBV核心抗原或HBV表面抗原。
9.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述HCV的抗原為E1、E2或NS3/4a抗原。
10.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述HIV的抗原為Gag(p55gag)、Pol、Vif、Vpr、Tat、Rev、Vpu、Env或Nef抗原。
11.根據實施方式10所述的疫苗藥劑學組合物,其特征在于,上述HIV抗原為Env抗原。
12.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述流感病毒的抗原為包膜糖蛋白HA或NA。
13.根據實施方式12所述的疫苗藥劑學組合物,其特征在于,上述流感病毒的抗原為包膜糖蛋白HA。
14.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述HSV的抗原為HSV的糖蛋白gB、gD或gH。
15.根據實施方式14所述的疫苗藥劑學組合物,其特征在于,上述HSV的抗原為HSV的糖蛋白gB或gD。
16.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述Hib的抗原為PRP或其共軛物。
17.根據實施方式16所述的疫苗藥劑學組合物,其特征在于,上述Hib的抗原為PRP和破傷風毒素的共軛物。
18.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述腦膜炎奈瑟菌的抗原為PI、PII、PIII、菌毛蛋白、脂多糖、鐵結合蛋白或蛋白酶體。
19.根據實施方式18所述的疫苗藥劑學組合物,其特征在于,上述腦膜炎奈瑟菌的抗原為蛋白酶體。
20.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述白喉棒狀桿菌的抗原為白喉毒素。
21.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述百日咳桿菌的抗原為百日咳毒素。
22.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述破傷風梭菌的抗原為破傷風毒素。
23.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述水痘病毒的抗原為活減毒水痘病毒、惰性減毒水痘病毒、gpI或gpII。
24.根據實施方式23所述的疫苗藥劑學組合物,其特征在于,上述水痘病毒的抗原為活減毒水痘病毒。
25.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述非毒性脂寡糖是對脂多糖進行堿處理,來實現脂質A的脫酰基化而被非毒性化。
26.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述非毒性脂寡糖具有1500~10000Da的分子量。
27.根據實施方式26所述的疫苗藥劑學組合物,其特征在于,上述非毒性脂寡糖具有2000~4000Da的分子量。
28.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,上述非毒性脂寡糖由腸埃希氏菌衍生。
29.根據實施方式1所述的疫苗藥劑學組合物,其特征在于,
上述藥劑學組合物還包含免疫佐劑;
上述免疫佐劑是選自包含Mg、Ca、Sr、Ba及Ra的組中的第二族元素、選自包含Ti、Zr、Hf及Rf的組中的第四族元素或鋁的鹽或其水合物。
30.一種樹突狀細胞的成熟化誘導用組合物,其特征在于,包含脫酰基化非毒性脂寡糖。
31.根據實施方式30所述的樹突狀細胞的成熟化誘導用組合物,其特征在于,上述非毒性脂寡糖是對脂多糖進行堿處理,來實現脂質A的脫酰基化二被非毒性化。
32.根據實施方式30所述的樹突狀細胞的成熟化誘導用組合物,其特征在于,上述非毒性脂寡糖具有1500~10000Da的分子量。
33.根據實施方式32所述的樹突狀細胞的成熟化誘導用組合物,其特征在于,上述非毒性脂寡糖具有2000~4000Da的分子量。
34.根據實施方式30所述的樹突狀細胞的成熟化誘導用組合物,其特征在于,上述非毒性脂寡糖由腸埃希氏菌衍生。
- 還沒有人留言評論。精彩留言會獲得點贊!